|
Russel Wallace : Alfred Russell Wallace (sic) Distribution of Wild Life in Time and Place (S684a: 1910)
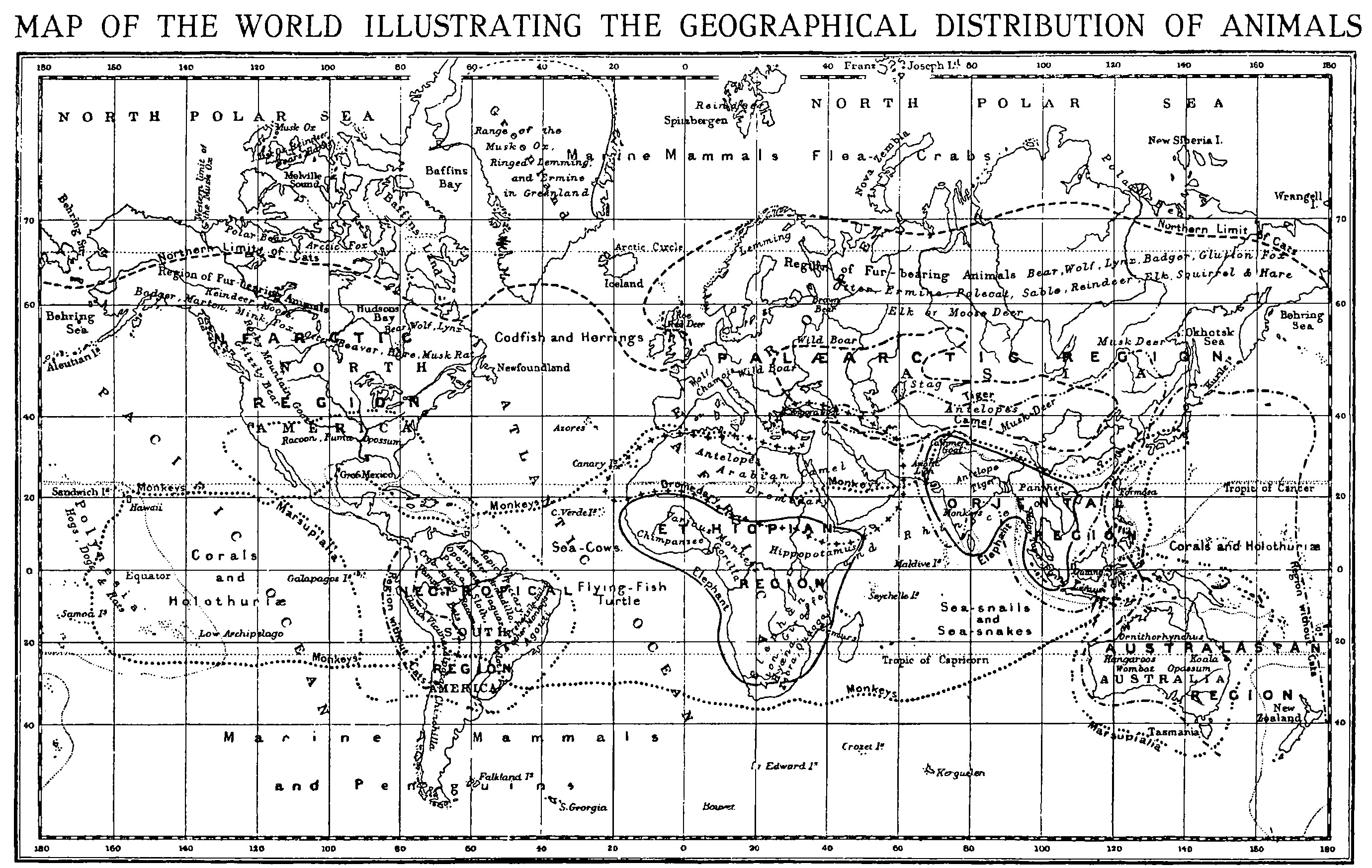
Darwin himself laid the foundations of the study by his observations and deductions as to the total absence of both mammalia and amphibia from oceanic islands, and also by his extensive experiments and observations on the modes of dispersal of plants and animals; and it is only by combining all these sources of information with those of recent inquirers as to the nature of the deposits in the deep oceans that any intelligible view of the subject has been arrived at. It may be as well first to state briefly some of the curious facts and apparent anomalies that have to be explained. Why, for instance, are deer entirely absent from Africa--except on the southern shores of the Mediterranean--though found over the whole of the other great continents from Patagonia to Greenland and from Britain to Japan? Why are camels found only in the great desert-belt of North Africa and Central Asia, and their nearest allies, the llamas and alpacas, only in the Andes south of the Equator and in Patagonia? The old idea that animals are found where the conditions are fitted for them is disproved by the fact that the entire horse tribe is now limited, in a wild state, to Africa and Asia, but when introduced into both North and South America by the Spanish conquerors they ran wild and increased enormously. Most extraordinary is the case of the curious tapirs, animals something like small rudimentary elephants, of which two or three species are found in Tropical America, and the only other species--the Malayan tapir--in Borneo and the Malay Peninsula. Among birds we have the humming-birds swarming all over America from Patagonia to Labrador and Alaska, but in no other part of the world. The equally wonderful birds of paradise, of which about fifty different species are now known, are found only in New Guinea and a few adjacent islands. Lastly, we have the great mammalian order of Marsupials, limited to Australia and the adjacent islands as far as Celebes and the Moluccas on the one hand, while one family of the same order, the opossums, abounds in South America and as far north as California and Virginia. We may also look at these differences from another point of view--that of the similarity of the animals in countries far apart, and the diversity that is often found between those which are comparatively near together. If a person travels by any of the ordinary routes from England to Japan or the Amur, he will have gone very nearly half round the globe; yet when he reaches those remote countries he finds in the forms of life a wonderful similarity to those of his native land. The fields and woods are tenanted by titmice, hedge-warblers, wrens, wagtails, larks, red-breasts, thrushes, and buntings, some absolutely identical with his feathered friends, others so nearly resembling them that it needs a close examination to detect the difference between them. Even among the insects he will notice many butterflies, beetles, and so on, that will be familiar to him, though they may prove to be distinct species. In the Amur valley he will find even the mammals to be very similar. The fox, the badger, the weasel, the otter, the roedeer, the hedgehog, the mole, the squirrel, and the Irish hare, are all the very same as the British species; while the wolf and the bear are identical with those that inhabited the British Isles within the historical period, and are still common in Europe. But if a settler in Australia goes to New Zealand, a distance of about 1,300 miles, he finds himself in a country zoologically and botanically entirely unlike the country he has left. Kangaroos, wombats, and the other varied forms of marsupials are absolutely wanting. There are no wild mammalia except two [[p. 58]] kinds of bats; the birds are almost all quite new to him; while insects are very scarce and altogether unlike the many handsome or curious species found in his own country. Even more remarkable, perhaps, is the case of two small islands, Bali and Lombok, in the Malay Archipelago, only fifteen miles apart, yet strangely different in their productions. Their birds differ more than do those of Britain and Japan. Bali has red and green woodpeckers, barbets, weaver-birds, black-and-white birds like large wagtails, and many others quite unknown in Lombok, where we find screaming cockatoos and friar-birds with bald heads, and the strange mound-building megapodes, all allied to Australian species and entirely absent from both Bali and Java. Again, the great island of Borneo is much more like Sumatra and the Malay Peninsula, both in its mammals and birds, than is the much nearer island of Celebes, whose productions are more nearly allied to those of India or Africa. Now, these and most of the other peculiarities in the distribution of animals are either directly explained or rendered intelligible by the application of the facts and general principles already enumerated; and I therefore propose now to explain as clearly and simply as possible the nature of those principles, which, when once thoroughly understood, will afford the means of solving almost all the problems of distribution both of plants and animals. PERMANENCE OF OCEANIC AND CONTINENTAL AREAS. I begin with what is perhaps the most fundamental and far-reaching of the phenomena on which the interpretation of the existing distribution of plants and animals depends, the true importance of which was first perceived by Darwin. Before the "Origin of Species" appeared, the common belief was that almost all the islands scattered over the great oceans were the remnants of former continents which had sunk beneath the waters, while our existing continents rose up to take their place. This view was accepted almost as a matter of course, and seemed, at first sight, to be quite in accordance with the fact that most of the older rocks in all parts of the world were of marine origin. But, during the voyage of the Beagle, Darwin was able to visit many of these islands, and he was struck by the poverty of their forms of life. He also noticed the fact that all of them were either volcanic or of coralline formation, and they never contained any of the stratified rocks characteristic of continents and of islands that have once formed parts of them. He therefore concluded that they had all been formed in the ocean itself, though some of them appeared to be very ancient; and the total absence of native land mammalia could thus be easily explained, as these animals, though often good swimmers, could never cross wide seas or oceans. He further showed by experiment that the eggs of batrachians (frogs, newts, etc.), which were equally wanting, were quickly killed by salt-water. Some years later, when electric cables began to be laid across the oceans in various directions, and when the voyage of the Challenger showed us the contour of the ocean floor and the nature of the deposits forming upon it, a striking confirmation of Darwin's views was obtained. For it was then for the first time shown that the floor of the great oceans did not form hills and valleys and mountain ranges, as had often been assumed; but was really an enormous, slightly undulating plain at a depth of from ten to thirty thousand feet, out of which plain the continents and islands usually rose abruptly; so that the 100-fathom line closely approached their shores, and even the 1,000-fathom line only left a narrow belt around them, in no way altering their general outline. Most important consequences result from these facts. Sir John Murray, of the Challenger expedition, has calculated that the mean height of the land of the globe is 2,250 feet above sea-level, while the mean depth of the ocean is 13,860 feet below it. But the area of the land is little more than one-third that of the oceans, whence it follows that the bulk of the oceans in cubic miles is more than thirteen times that of the land. In my "Darwinism" (p. 345) I have given a diagram, showing at a glance these proportions; and I pointed out, I believe for the first time, that these great differences in the bulk of land and water rendered it almost impossible for any interchange of continents and oceans to have occurred during geological time. There must have been some approach to equality in the simultaneous rising and sinking of the solid crust of the globe. But if one of our smallest continents were to subside, and an adjacent portion of the ocean floor on either side of it to rise, the former would almost wholly disappear before the latter had approached within many thousand feet of the surface. And if a similar change went on successively, one [[p. 59]] continent after the other would sink, while the counter-balancing rise would hardly have rendered the vast extent of ocean any shallower than it was before. The contour of the ocean bed, and the comparative bulk of land and water, alone serve to indicate that the great and often-repeated vertical movements of the land over all the continents throughout geological time must have occurred within or on the margins of the areas they now occupy. If they had not been thus limited, if the gigantic forces of elevation and depression which have been always at work in the continents had been also at work beneath the ocean floors, the latter must have exhibited some of the abrupt inequalities of the former, instead of being entirely different from them in their contours. These conclusions are quite in accordance with the broad features of stratigraphical geology in every part of the world. Everywhere we find the successive strata of which the earth's crust is formed occurring in long, irregular, but more or less parallel belts overlapping each other; and in numerous cases exhibiting intercalated land surfaces or fresh-water deposits. This may be well seen by looking at any large geological map of England, or of any part of the Continent. This feature is perfectly explained by the minute examination that has now been made of the ocean floor near the land, as well as across its greatest depths. It has been found that the larger part of the débris of continents brought down by rivers is deposited mainly within a distance of from 50 to 100 miles from the coasts, while the very finest mud, which, of course, sinks more slowly, is deposited within 150 miles, and only in rare cases extends so far as 200 miles. All these are termed "shore deposits," and they form, when consolidated and uplifted, the successive strata of the earth's crust. But beyond these limits, and extending for thousands and tens of thousands of miles over the whole floor of the deep oceans, is found a very different set of deposits termed "oozes." The most extensive is that termed globigerina ooze, because it consists very largely of the shells of a minute organism of that name, and it is found at almost all depths from 250 to 2,000 fathoms. The remains of many other surface organisms occur among it, as well as of those that live at the bottom; but no true land débris. There are, however, numbers of volcanic particles, either derived from pumice, which floats for enormous distances, or from volcanic dust carried by the wind, which in the case of the Krakatoa eruption fell on the decks of ships nearly [[p. 60]] 1,000 miles from the volcano; while, what is still more extraordinary, a small proportion of meteoric dust can always be found. This gives an idea of the extreme slowness with which the oozes are formed, since such dust can hardly ever be detected on land. Another indication of the same thing is the large number of sharks' teeth, and the ear bones of whales, the only parts of these animals which escape complete disintegration. In these vast areas of deep-sea oozes covering the whole ocean floor we have another contributory proof that oceans and continents are not interchangeable. For if they had been so, as many writers still suppose, then each upheaved continent must rest on a basis formed of such consolidated oozes, with their characteristic fossils and inorganic particles. Yet in not a single one of our continents has any such base-rock been found, while our greatest geologists are almost unanimous in declaring that such consolidated oceanic oozes occur nowhere in the whole series of strata in any part of the world. It is a very suggestive fact, as indicating that the law of subsidence balancing elevation is a real one, that all the greatest depths yet found in the ocean are in the vicinity of lofty volcanoes or ranges of mountains. Thus a depth of 23,250 feet has been found a little north of Porto Rico, West Indies; 21,930 feet a little south of the Aleutian Islands; 27,930 feet a little north-east of Japan; 18,140 feet a little north of the Sandwich Islands; 19,000 feet a little north-west of Bermuda. Very few of these very deep soundings have, however, been taken, as they are exceedingly laborious and costly. The conclusion, therefore, of many of our greatest geologists, that the whole series of the stratified rocks present evidences of having been formed near to land, is supported by so many weighty arguments, derived from various sources, that it must be considered to be a well-established deduction of science. Yet we still find many palæontologists insisting on land connections across the deepest oceans in order to explain similarities in their extinct animals. But there are other ways of explaining such facts much more in accordance with probability, and with the great generalisation I have here indicated. EXISTING CONTINENTS AND THEIR PROBABLE EXTENSIONS. If we look at a globe or a map of the world in hemispheres, and consider the position of the continents and larger land masses, we find that they, even at the present time, are wonderfully connected; that it would be actually possible for a man to traverse the whole of the continents, starting from Cape Horn, without ever going out of sight of land or requiring any other vessel than a small open sailing-boat. If we now look at a map showing the 1,000-fathom line around all the continents (such as that given in my "Darwinism"), and taking this as roughly indicating the amount of extension of the land during the whole Tertiary period of geology, we shall find all the continents united towards the north by wide stretches of land, offering ample facilities for the migrations of land animals at successive epochs. WARMER CLIMATES IN PAST GEOLOGICAL PERIODS. But there is another consideration which is continually left out of account by those who still claim direct land connections to account for real or supposed affinities of the extinct animals of South America and South Africa, and that is the much warmer climate that prevailed in northern and Arctic regions throughout Tertiary times, and even very much earlier. This is proved by abundant remains of fossil plants found in Miocene strata all round and within the Arctic Circle, and of such a character as now grow a long way further south. On the west coast of Greenland, in 70° N., are beautifully preserved specimens of such trees as chestnuts, [[p. 61]] sassafras, oaks, planes, beeches, plums, vines, and even a magnolia, all closely resembling trees and shrubs which now grow 20° or 30° further south in North America. Even so far north as Spitsbergen, one of the most barren and most inhospitable regions on the globe, a rich fossil flora has been found, indicating a climate fully as mild as that of the warmer parts of Canada at the present time, comprising such familiar plants as hazel, ash, and walnut, with water-lilies and an iris. As there was probably continuous land between Europe, America, and Asia, at least as low as the latitude of Stockholm, while the land a little further south had a warm temperate climate, it became possible for every kind of mammal of the temperate zones and many of the Tropics to migrate from one continent to the other. There is also evidence that still warmer climates prevailed in the Cretaceous period, and, therefore, almost certainly in the early Tertiary, when all the chief types of mammalian life were well developed. In strata of this period in Greenland, not far from those of Miocene age above referred to, not only many of the trees of the later period occur, but, intermingled with them, such indications of warmer conditions as figs, myrtles, cycads, and tree-ferns. The only justification for postulating direct communication is that a shorter route is more effective, and therefore more probable, than a longer and more circuitous one. But this argument supposes two things which no geologist will admit--first, that the geological record itself is at all near to a full and continuous one; and, secondly, that our knowledge of the actual record approaches to completeness. It is, on the contrary, held to be certain that the record of the animal life of all former periods contained in the rocks is extremely imperfect and fragmentary, and that our knowledge of what exists, much of it buried deep in the crust of the earth, is wholly inadequate. It is interesting to note that the very changes in the height of land and elevations of sea-bottoms in the northern parts of the Atlantic and Pacific oceans, which would facilitate the migration of land animals between the eastern and western continents, would themselves greatly assist in the production of the warm climates we find proofs of. It is universally admitted that warm ocean currents, aided by the warm winds that carry moisture from them, are the great sources of the mild and equable climates that even now prevail in many parts of the north temperate zone. The British Isles, as well as Norway, owe their mild winters to the Gulf Stream and the warm and moist westerly winds that are so prevalent; while Labrador, in the same latitude, has a thoroughly Arctic climate, due to its being washed by a Polar current, and being exposed to cold winds from the Arctic regions of America. But when the North Atlantic was elevated about 1,000 fathoms, the heating power of the Gulf Stream would be greatly augmented by two causes. It would be more concentrated by being kept out of the Polar seas, and would circulate more rapidly around the northern shores of the Atlantic. It would also be completely free from the Arctic icebergs which now pour into it as soon as the ice breaks up in the Polar basin, causing that wintry weather which is regularly experienced in Great Britain during the first half of May. And when such a mild climate prevailed as is shown by the abundant tree and shrub vegetation within ten degrees of the Pole, there would probably have been very small accumulations of ice, which would have no more serious effect on the climate than have the glaciers of Switzerland or of New Zealand in their respective cases. [[p. 62]] EVIDENCE OF STILL EARLIER WARM ARCTIC CLIMATES. Before quitting this part of our subject, it will be well to note the abounding evidence of equally warm climates in the northern hemisphere throughout much, if not all, the Secondary period of geology. I will first notice the suggestive fact that, even more recently than the Cretaceous beds of Greenland above mentioned, in the London Clay (Eocene) of Sheppey, Bournemouth, and a few other localities, abundance of fossil plants has been found closely allied to living tropical or sub-tropical forms. Fruits of palms allied to those of the Nipa of Indian and Bornean river swamps are frequent, as well as the fruits of a proteaceous plant whose allies are now found in Australia. There are also species of acacia and anona, both warm-country forms; and, associated with these are found teeth and bones of crocodiles, a large sea-snake, and several mammals of the hog and tapir tribes. There are also numbers of marine shells now characteristic of tropical seas, including the nautilus. Throughout the Secondary period similar conditions appear to have prevailed, the sea-shells being, both in size and structure, almost wholly of the type of those now confined to the Tropics. The abundance of large and often gigantic reptiles--terrestrial, marine, and aerial--which are found in these strata throughout Europe and America are equally indicative of tropical conditions. And these conditions must have prevailed also within the Arctic Circle, since the very early Triassic strata of the Parry Islands, in 77° N., contain many ammonites as well as the vertebræ of the large marine reptile ichthyosaurus; while, in the Triassic beds of Spitsbergen, ammonites and nautilus are found, and still earlier in the Coal Measures of that country are lepidodendrons and large fronded ferns. We thus see that during the whole range of geological time between the Middle Tertiary and the Carboniferous ages there is almost continuous evidence of such mild and uniform climates far into what are now Arctic lands as would suffice for the temporary residence and continuous migration of most tropical animals between the Eastern and Western continents. No adequate reason, therefore, can be given for assuming an enormously difficult if not impossible bridging over of the great oceans, in order to account for the existing distribution of animal life; and still less in order to explain its apparent distribution in remote geological ages, as to the whole conditions of which we have such very scanty and imperfect knowledge. Even now many wholly tropical animals live very well and breed freely in the whole temperate zone, such as domestic fowls and peafowl; while monkeys are found as high as 11,000 feet in parts of the Himalaya. As late as the Pliocene period they lived in France and Britain. Even the hippopotamus, an almost tropical animal now, inhabited South Europe, and ranged as far north as Yorkshire during the summers of the Glacial period! THE POWERS OF DISPERSAL OF ANIMALS. The distribution of animals over the earth's surface of course depends in part upon their individual powers of locomotion--on land, through the waters, or through the air--but this is by no means the only factor of importance in determining their actual habitations, and is often entirely neutralised by other causes. In most continents, and in many of the larger countries or islands, birds exhibit a very similar restricted range to mammals, notwithstanding their very superior powers of locomotion. Reptiles also exhibit little difference in this respect, except that they are more dependent on temperature and become scarce in temperate and almost absent from very cold countries. The amphibia--frogs, toads, newts, salamanders, etc.--are in some respects more restricted than reptiles in their ranges, and more extended in others. This is due to the fact that they can withstand a lower temperature, their eggs being often frozen without injury; but, on the other hand, they are killed by salt water, and this explains the interesting fact that they, as well as mammals, are wholly absent from all true oceanic islands. This class of islands may be defined as being situated in the great oceans far from any extensive land masses, and being surrounded by depths of more than a thousand fathoms. All such islands, as was first pointed out by Darwin, are entirely without any of the older stratified rocks, consisting either of volcanic rocks or of coral formations which have been deposited upon them. Such are the Azores, Madeira, St. Helena, Mauritius, and all the remoter islands of the Pacific Ocean, none of which possesses either mammalia or frogs, and the most remote from land no reptiles or freshwater fishes. These are entirely absent from the Azores and St. Helena, while the Sandwich Islands possess two lizards--one very widespread over the whole western Pacific, and therefore presumably conveyed accidentally in canoes; the other a peculiar gecko, whose ancestors were probably introduced at some remote epoch by exceptionally favourable circumstances, such as large floating trees from more westerly islands. Within the Tropics especially, masses of trees and floating vegetation are often carried out to sea, and under favourable conditions may be driven by winds and currents for many hundreds or even thousands of miles, and carry with them small lizards or their eggs, insects, and land shells. These latter often hide in crevices or under bark, while some snails, when dormant, will stand immersion in salt water for twenty days without injury. We thus see an explanation of the curious fact that in the remotest islands, even when every kind of vertebrate except birds is absent, a few land shells and terrestrial insects, especially beetles, are always to be found. But the more remote the island, and therefore the rarer and more exceptional the chances that bring these latter creatures to it, the more peculiar we find the existing species to be. Thus, in the very remote Sandwich Islands, with their rich vegetation and favourable climate, there are about 500 different kinds of land shells, almost the whole of which are peculiar species; and the moderately rich insect fauna is equally peculiar. In the Azores, however, [[p. 64]] which are less remote from the nearest continent, and much more subject to violent storms--bringing numerous birds which are both seed and shell carriers--shells and insects are less numerous, and a much larger proportion belong to European forms. All the facts connected with these oceanic islands, some of which form groups of great extent and of considerable geological age, and are scattered over all the great oceans, strongly support the view that the oceans themselves are permanent features of the earth's surface; while the complementary fact, that the great continents in their main features and position are equally permanent, is demonstrated, first, by the occurrence in all of them of an almost identical series of stratified rocks of all the geological periods, and, secondly, because these rocks everywhere show plain indications of having been deposited very near the margins of the lands from the denudation of which they have been built up. With the amount of preliminary knowledge here sketched out, we are in a position to explain most of those curious examples of discontinuous distribution which often appear so puzzling. THE DISTRIBUTION OF TAPIRS. One of the most prominent of the puzzles of distribution to the earlier naturalists was that of the tapirs, the two commonest species of which inhabit Tropical South America from Brazil to Paraguay, and the Malay Peninsula, Sumatra and Borneo. Three other species also inhabit Tropical America, one in the high Andes and two in Central America from Panama to Mexico; but none is found either in Africa or continental India. But the course of geological discovery during the nineteenth century has completely explained this apparent anomaly. First, in 1825, Cuvier described the skeleton of the Palæotherium from the early Tertiary beds of Paris as being allied to the living tapirs; and later, both in France and Germany, the remains of true tapirs were found in the middle Tertiary (Miocene) and late Tertiary (Pliocene) strata, both in France and England. Fossil tapirs have also been found in China and in North America from Carolina to California. Now, always keeping in mind the extreme imperfection of the geological record, these discoveries clearly indicate that many species of tapirs inhabited warm and temperate Asia, Europe, and North America in middle and late Tertiary times. But during these epochs warm and even sub-tropical conditions prevailed in the Northern hemisphere as far as the Arctic Circle, accompanied by an abundant and luxuriant vegetation; and we have also seen that these conditions were probably brought about, in part, by a greater extension of land in the North Atlantic, admitting of more or less easy communication between the Eastern and Western hemispheres. The problem of the tapirs is, therefore, completely solved. DISTRIBUTION OF THE CAMEL TRIBE. The case of the camel is perhaps even more interesting than that of the tapirs, which it somewhat resembles. Camels are now inhabitants of the desert regions of Western Asia and North Africa, and of no other parts of the world. But in the Andes and temperate plains of South America is a group of small animals--the llamas, alpacas, and guanacos--which in all essentials of structure, even to the peculiar complex stomach adapted for water-storage, belong to the camel tribe. These two allied groups are, therefore, now almost as remote from each other as are the American and Malayan tapirs, with no allies whatever in intervening regions. But here, too, geology has furnished the solution. First, in the late Tertiary deposits of the Siwalik hills of North-west India remains of true camels have been found; but, during the last thirty or forty years, a wonderful series of fossils has been discovered in North America, in late, middle, and early Tertiary strata, by which the whole gradual development of camels from a smaller and more primitive type has been exhibited. The same strata have shown us early forms combining the characters of all the hoofed animals, from the swine and the hippopotamus to camels, cattle, deer, sheep, and antelopes, gradually becoming specialised into those varied forms. It is, therefore, clear that in all probability the camel and llama tribes originated in the Central United States, where, towards the end of the Tertiary period, they became extinct. Previous to this catastrophe, however, some of the true camels migrated to the Eastern hemisphere, probably by the way of continuous land in the North Pacific, and have left as their only survivors the camel and the dromedary. About the same time, and probably driven to migrate by the same adverse conditions which led to the extinction of so many of their allies, the llama group passed southwards along the central mountain ranges into South America, where they have found suitable conditions for their survival south of the Equator, in the high Andes, and on the arid plains of Patagonia. Here, again, the problem is satisfactorily solved. THE DISTRIBUTION OF THE HORSE TRIBE. The horses, asses, and zebras constitute a distinct and very remarkable tribe of mammals, being the only members of that great class which possess a single functional toe on all four feet, forming the well-known horse's hoofs. These animals are now strictly limited in a wild state to Africa and Western Asia, though in quite recent times they ranged over Europe and the British Islands; and many allied groups are found in still earlier times, back to the early Tertiary, showing the gradual transformation of a four-toed animal of small size, step by step, into the true one-toed horse. But, what is more curious still, a similar and even more complete series of ancestral forms existed in North America, where the true horse was also developed, its fossilised remains being found over both North and South America nearly up to the Glacial period. Then suddenly all became extinct. But after the great Ice Age had come and gone, with the many changes of animal life that accompanied it, the two continents again became well adapted [[p. 65]] to horse life, as proved by the fact that, as we have said, when introduced by the early Spanish conquerors they ran wild, and increased enormously both in the western plains of the north, and in the llanos and pampas of the south. It is curious that the series of these ancestral horses runs almost parallel in America and Europe, though the species are not exactly the same; and this seems to imply that the North Atlantic land connection existed for a long period, and that intercommunication from America to Europe and from Europe to America frequently occurred. It is, in fact, difficult to tell in which continent the true horses now living actually originated. In both there was an animal about the size of a fox which seems to be the earliest direct ancestor of the modern horses and asses. This intermingling of the ancestral forms of a group so well adapted for migration both in structure and habits renders it certain that such connecting lands and such mild climates as are indicated by other evidence really existed throughout a considerable part of the Tertiary period. This strange history has been made use of by a talented American writer, Mrs. Charlotte Perkins Gilman, to point a moral against those who oppose the possibility of the social advancement of humanity. It is one of three such illustrations in a poem under the heading of "Similar Cases" in a volume entitled "In This Our World." The name "Eohippus" was given by Professor O. Marsh to one of the less-known remoter ancestors of the horse. "SIMILAR CASES." There was once a little animal Said the little Eohippus: Then they laughed enormous laughter, They considered him disposed of, ELEPHANTS AND THEIR ANCESTORS. The elephants are of special interest for two reasons. Their distribution in middle and late Tertiary times was very similar to that of the horses, and they disappeared from America, Europe, and Northern Asia at as late or even later a period and with equal suddenness. The huge mammoth with its enormous curved tusks lived in Northern Europe and Asia as well as in North America down to the human period. Many other species are of late Tertiary date and died out a little earlier; and among these were some curious small elephants from three to five feet high, whose remains are found in the caves of Malta and Cyprus. The allied mastodons have a similar distribution at a [[p. 66]] somewhat earlier period, and these ranged over both North and South America and survived there till the appearance of man. None of these numerous forms gives any clue to the early stages of the elephant type so strikingly different from all other mammalia. But within the last ten years remarkable discoveries have been made, in the early Tertiary beds of the Fayum valley in Middle Egypt, of an interesting group of fossils of mammals which are believed to be the ancestral forms of the entire elephant tribe. The earliest of these is an animal not very different from the ancestral swine and tapirs, and of very small size. The whole series is not yet complete, but the following short account, from the "Guide to Fossil Mammals" in the British Museum, very clearly explains its bearing upon the problem. "The fossils, so far as known, show that the earliest forerunners of the elephants were small marsh-dwellers which lived on a succulent food in the African region. They gradually increased in size without essentially altering their limbs or bodies; but as their legs lengthened and their neck shortened, their face and chin gradually became elongated to reach the ground for browsing. When this strange adaptation had reached its maximum degree, the chin shrivelled, leaving the flexible toothless face without any support. Thus arose the unique proboscis of the elephants, which has become prehensile by stages which cannot be traced, because soft parts are not preserved in ordinary geological formations." In the "Guide to Recent and Fossil Elephants," 1908, there is a diagram showing the proportionate sizes of the three chief types of fossil elephants previously known, together with the two recent discoveries in Egypt, which is very instructive. In these we see the progressive shortening of the chin, the straightening of the tusks, then their bending downwards instead of upwards, and lastly their reduction to ordinary-looking teeth in both jaws. It is also very suggestive that this change occurred at a period when Africa was cut off from the rest of the world by a sea extending from the Atlantic across Northern India. These early forms were thus subject to special conditions from which they could not escape by migration, and thus gave full play and ample time for the action of variation and natural selection to initiate the main elephantine characteristics. When, in the middle Tertiary, free land connections were formed to Asia, Europe, and North America, elephants had attained such a size and such completeness of organisation that they were enabled to spread over the two hemispheres, becoming modified in each into special forms adapted to the new conditions. Then, after a long period of success, a number of competing tribes arose, such as the extensive and varied hoofed ungulates culminating in the cattle, deer, and antelopes, as well as a host of large and powerful carnivores culminating in the great sabre-toothed tiger, when the elephants everywhere became extinct, except in the two most favourable localities for their maintenance--that is to say, in the equatorial forests of Africa and Asia. WHY ARE DEER AND BEARS ABSENT FROM TROPICAL AND SOUTH AFRICA? The Tropical African region affords us one of the most interesting problems in zoological geography. In many of its larger mammals and birds it presents a considerable resemblance to those of Tropical Asia, yet there are some most remarkable deficiencies. The most singular is the absence of the whole of the deer and the bear tribes, which are found in all the other continents, together with the camels, goats, and sheep which abound in the adjacent parts of Asia. Now, the explanation of these peculiarities is to be found, first, in the existing fauna of the great island of Madagascar; and secondly, in that isolation of Tropical and South Africa from the rest of the world during the early portion of the Tertiary period, which is proved by a vast extent of early Tertiary marine strata to the north of it, and which has been already mentioned as having led to the early development of the elephants. The interest of Madagascar lies in the fact that the evidence all points to its having been formerly united with Africa, but only at a time when that continent possessed a much more restricted fauna than now. For although Madagascar contains about seventy different species of mammals, these are mostly of low type, while none of the larger and more highly developed African groups is found there. Though there are nearly forty of the lower quadrumana (the lemurs), there is not a single monkey or ape. The great carnivora of Africa--the lions and leopards, the hyænas and lynxes--are absent and are represented only by eight civets, all of peculiar genera; the very numerous rodents of Africa, including squirrels, porcupines, and hares, are absent, the order being represented by a few peculiar mice; while the ever-present antelopes of Africa--nearly 100 species--as well as its zebras, rhinoceroses, and elephants, are quite unknown. In place of all these we find only a river-hog, while a small hippopotamus seems to have recently died out, both of which, being semi-aquatic in habits, might, in very favourable circumstances, have crossed the Mozambique Channel. The reptiles show similar singularities, most of the genera being peculiar; while two families of snakes, both abundant in Africa, are wholly wanting. Even the birds exhibit the same phenomena. The genera and some of the families are quite peculiar, but they seem to be related to Indian as much as to African types. A number of specially African birds, such as hornbills, plantain-eaters, and barbets, are absent. We thus find that all the facts go to prove that the fauna of Africa must have been very poor as regards variety of its higher animals at the time when Madagascar was separated from it; and that epoch is demonstrated to be a rather remote one by the distinctness of the two faunas now, and also by the number of peculiar species and genera of the higher mammalia and birds characterising that continent. We conclude, therefore, that the separation from Africa dates from the early Tertiary period. For the later history of animal life in Africa we are indebted to the geological record in other continents. We find that all the larger and more [[p. 67]] characteristic of the great mammalia of Africa inhabited Southern Europe and Northern India in the middle and late Tertiary periods. Such are the great carnivora--lions, tigers, and hyænas, the extensive and varied hoofed animals, from the zebras, rhinoceroses and swine, to the antelopes, the giraffes, and even the okapi--all of which have left their bones or skeletons in middle or late Tertiary strata. It becomes clear, therefore, that not long, geologically speaking, after Madagascar was separated from Africa, the Saharan sea was partially elevated so as to open one or more free passages into the African continent. Then the highly organised mammals just enumerated would migrate into the new and inadequately peopled land, would there increase rapidly, and develop into a variety of distinct species fully adapted to the new conditions. Then came the third and last great change, the simultaneous subsidence of the Mediterranean area, and the elevation of that of the Sahara and of Arabia, bringing about the desert conditions of these countries, and thus isolating Tropical and South [[p. 68]] Africa. Thenceforth the typical African fauna was left to develop uninfluenced by immigrations from the extensive Euro-Asiatic continent, leading to that endless variety and abundance of "great game" in which this region is pre-eminent. And now, at the end of this strange series of geographical changes and zoological migrations, we find the solution of the problem of those deficiencies which are so remarkable a feature of this continent. For the varied and widespread deer tribe only reached their full development in the later Tertiary period; while the true bears seem to have originated about the same epoch, and to be in fact at their fullest development in our own time. These two groups, therefore, either did not exist in the adjacent regions, or were so comparatively weak in structure as not to be fitted to take part in the great rush of migration to the newly opened land. The absence of camels is probably due to the fact that, as already shown, they originated in North America and entered Asia from the north-east. They had reached North-west India towards the latter part of the Tertiary period, and were too remote from Africa to take part in the great incursion. The goats and sheep seem to be among the very latest developments of the large tribe of Bovidæ, or hollow-horned ruminants, and as they are pre-eminently mountain animals, they would not have been in a position to take part in a low-land migration, even had they been in existence at the time, which they probably were not. As the whole subject is both vast and complex, I will here re-state the fundamental facts and conclusions on which a true explanation of all cases of anomalous distribution must be based. (1) The most fundamental of all is the proof, by the agreement of several distinct lines of argument, that the great continental and oceanic areas are not interchangeable, but are permanent features of the earth's surface, subject only to marginal extensions and reductions within fairly well-defined limits. (2) The slow but important changes of climate that appear to have been always going on. The more important of these for our special inquiry is the proof of an almost tropical climate in much of the temperate and sub-Arctic zone in the early Tertiary, changing gradually to warm-temperate towards its close, and then merging into the oncoming Glacial period. This must never be overlooked, as it often is, because, with moderate land extensions in the northern parts of both the Atlantic and Pacific Oceans, it offers ample means of migration, even of warm-country animals, between the several continents, and entirely obviates the supposed necessity for repeated bridging of the tropical and sub-tropical oceans, which has been shown to be unsupported by any evidence, opposed to a large body of facts, and almost, if not quite, physically impossible. Other factors of secondary importance are: the various powers and facilities of dispersal by the different groups of animals; the evidence of geology as to the preceding forms of life in the same areas and elsewhere; the frequent changes of climate and the nature of the vegetation; lastly, the continuous action of the competition of better-adapted forms of life, rendering it impossible for newcomers to maintain their existence, even if occasionally introduced into a new country. We thus see that the present distribution of the whole vast complex of living organisms is the final outcome of the entire course of evolution from the earliest dawn of life. The locality and range of every animal and plant may therefore be looked upon as one of its vital characters. It is what it is and where it is because it has been developed in close adaptation to the physical and organic environment in which it lives, and where alone, except in very rare cases, it is able to maintain its existence. The higher and more completely adapted forms--the dominant and aggressive species, as Darwin termed them--will always be able to maintain themselves, sometimes at the expense of native forms, when they gain access to a country previously occupied by lower types, as was the case when the highly developed animals of the Euro-Asiatic continent gained access to Tropical Africa. We see the same thing occur in the case of the European sparrow introduced into North America and the common rabbit into Australia, both of which have become pests in their new homes. But they have in both cases been assisted by man's interference with the native fauna and flora, and it is very doubtful if the same results would have followed any chance introduction of these species into countries where man had not already interfered with the balance of competing life. THE DIVISION OF THE GLOBE INTO ZOOLOGICAL REGIONS. In order to describe and compare the distribution of the various species, genera, and families of animals, it has been found necessary to mark out a certain number of extensive areas characterised by distinctive forms of animal life. The older naturalists usually adopted the great geographical and racial divisions--Europe, Asia, Africa, America (North and South), and Australia, or the still broader Arctic, Temperate, and Tropical Regions. They also used the general term "India or the Indies" for all Eastern tropical lands, sometimes including all the Tropics, as in our still common but misleading term "West Indies." The first thoroughly scientific attempt to establish a series of regions that should accurately summarise the main facts for any extensive class of animals was made by Dr. P. L. Sclater in 1857 for the class of birds, at that time, as it is now, probably the best-known of all the more extensive groups of animals. He was the first to point out that Europe and Asia do not correspond to primary divisions of animal life, as shown by the striking similarity and considerable identity of both the birds and mammals across the whole of Europe and temperate Asia. This formed his first great region, which he termed the "Palæarctic," as including all Old World northern lands. Then came the tropical portion of Asia, which possessed hosts of altogether peculiar species, genera, and whole families of birds not known in the temperate zone. This he termed the "Indian Region," because India in a wide sense
[[p. 70]] formed the bulk of it. This term in my "Geographical Distribution of Animals" I altered to "Oriental Region"--perhaps unnecessarily--because it included Burma, Siam, and all the great Malay islands, and this name has been widely adopted. It may be mentioned that the Palæarctic Region also included North Africa as far as the Sahara, all its chief productions, both animals and plants, being closely allied to those of Europe or Western Asia. The remainder of Africa, possessing a large proportion of peculiar types, and being thoroughly isolated from the rest of the world, constituted the "Ethiopian Region." The fourth was the "Australian Region," perfectly characterised by its very peculiar marsupial mammals, as well as an immense number of peculiar genera, and of several remarkable families of birds. The eastern half of the Malay Archipelago belongs to it, as do also New Zealand and most of the Pacific islands. Then came South America, including the West Indies and Central America as far as Mexico, forming the "Neotropical Region," because it included the whole tropics of the New World. Temperate and Arctic North America constitute the "Nearctic Region" of Dr. Sclater. It has relations with both the Neotropical and Palæarctic regions, but has sufficient special features to be kept distinct. It will be seen that the past geographical history of the several regions, as we have been able to trace it both by stratigraphical and palæontological evidence, enables us to understand why each of these regions possesses broad distinctive features while presenting certain resemblances to several others. By laying undue stress on these resemblances the primary zoological regions have been reduced in number and altered in constitution by some writers, so as to render them of little use or interest to the general reader, or even to the general student of zoology. Professor Huxley, for example, proposed a threefold division--Arctogæa, including the whole Northern hemisphere, with Africa and Malaysia; Notogæa, for South America and Australia; while New Zealand formed a region by itself. This division was founded on the prevalence of a low type of birds and of mammals in Notogæa, and on the lowest existing reptile--the curious lizard-like hatteria, found only in New Zealand, and there almost extinct. But these all depend on features of anatomical structure only appreciated by specialists, while a single region--including Europe, Asia, Africa, and North America--is absolutely useless for any practical purpose. Mr. Andrew Murray united India and Africa as one region, and North and South America as another; while several modern writers insist that North America is too much like Europe regions and North Asia to be separated from them, and thus form what they term the "Holarctic Region." As this last is rather widely adopted owing to its having been advocated and taught at Cambridge for many years by Professor Alfred Newton, especially as regards the class of birds, I will give a few of the main facts which show how great and fundamental are the differences of the two regions. No fewer than twelve out of the thirty-six families of land birds inhabiting the Palæarctic region are not found in the Nearctic; which latter has eight families not in the Palæarctic. Of genera, there are 118 which are Palæarctic but not Nearctic, and 113 Nearctic but not Palæarctic. Stating some of these facts in popular language, we find in North America three very common families of birds--the wood warblers (Mniotiltidæ), the hang-nests (Icteridæ), and the tyrant birds (Tyrannidæ)--containing together nearly 100 species spread over the whole country, all three of which families are totally unknown out of the American continent. In Europe and North Asia we have the hedge warblers (Accentoridæ), the fly-catchers (Muscicapidæ), and the starlings (Sturnidæ), quite unknown in America; besides more than twenty genera with over 100 species of warblers and their allies, as well as twelve genera with fifty species of pheasants and their allies, all equally unknown in America. Summarising the whole, we see that, in either region, the genera of birds which it has in common with the other do not amount to one-third of the whole number it possesses; while the species common to both, mostly Arctic, only amount to twenty. So far as the mammalia are concerned, there is a rather larger amount of resemblance, but even in their case the diversity is still ample. But, further, when we consider the very important part North America has taken in the development of the higher animals, especially of the great tribe of ruminant mammalia, including such specialised types as the camels and the horses, which seem to have been peculiarly its own, any division of the earth into zoological regions which obscures or ignores this past history by uniting this vast country either with Eurasia or with South America to form a single region must be equally inadmissible. The same objection applies to uniting Africa with India. Each has its peculiar families, and a very large number of peculiar genera of all the higher animals; but, because these are often small and not generally known, the groups of larger and more conspicuous mammals and birds which are common to both are allowed to obscure the more fundamental diversities. Even if we do, against the weight of evidence, make any such compound regions, we must immediately separate them again as "sub-regions," or we lose all the advantage of easy and intelligible classification of the many peculiarities of the Ethiopian fauna in all its groups from mammals to insects. It is for these reasons, which I have given in full detail in my works on the subject, that I still thoroughly hold the scheme of six primary zoological regions proposed by Dr. Sclater to be in all respects more accordant with the great facts of the existing distribution of animals as well as with their geological history, and the past geographical changes of the earth, than any which has since been proposed. As the most accurate and most useful classification for the student, as well as the most intelligible and most illuminating for the general reader, it holds the field against all attempts hitherto made to improve it.
"As the most accurate and most useful classification for the student, as well as the most intelligible and most illuminating for the general reader, it holds the field against all attempts hitherto made to improve it." In this last sentence of his article "Geography of the Animal World: Distribution of Wild Life in Time and Place," Wallace is referring to Philip Lutley Sclater's 1857 classification of the world's animal life into six biogeographic regions. Wallace himself relied on Sclater's scheme as the backbone to his own grand work, The Geographical Distribution of Animals (1876). What is most striking about this article, written by Wallace in 1910 and intended for a lay audience, is well encapsulated by the closing sentence cited above. Reading an article that is now over 100 years old, I am struck by the completeness of thought and by how much of the 'big picture' of natural history was already complete at that time, by and large thanks to the work of naturalists like Wallace. The elements summarized in this article are the usual main ingredients of any modern natural history: evolution, geological time, and climatic change. Wallace uses these three basic concepts to explain numerous oddities of animal distribution in what is effectively modern scientific prose. It took the natural sciences a long time to develop and agree on the validity of three fundamental facts: biological evolution, an ancient Earth, and global climatic shifts. These three elements are today the foundation of any scientific explanation of patterns in natural history, whether in the ecological, zoological, or paleontological fields. Over the last 100 years we have come to document and understand the patterns and processes underlying evolution, geological time, and climatic change, knowing orders of magnitude more about these aspects of Earth history now than Wallace ever did. For example, Wallace goes to some length to defend the 'permanence of oceanic and continental areas,' when today we know that the configurations of the continents and oceanic basins are constantly changing (though the acceptance of continental drift itself took decades). To Wallace's reference to 'dominant and aggressive' and 'highly developed animals of the Euro-Asiatic continent' in relation to those of Africa, a modern reader cannot help but raise an eyebrow. Though certainly unintended by Wallace, a whiff of the 19th and early 20th century colonialist climate of the time does come through. One topic conspicuously not addressed in this article is the biogeographic history of humans. One hundred years on, through abundant genetic and paleontological data, we know today what naturalists in Wallace's time might only have guessed: all clues to our origins point firmly to Africa, with our dispersal and diversification across the other continents taking place only in the recent geological past. Thinkers like Wallace were major contributors to the highly contentious and exciting sea change in scientific thinking that took place in the late 19th Century. More than 100 years on, his writings hardly show their age. Reading a synthesis such as this article may be the best reminder that we modern practitioners of the natural sciences are very much the direct intellectual descendants of Wallace. |