|
Russel Wallace : Alfred Russell Wallace (sic) in the History of the Evolution Theory. (S389: 1886)
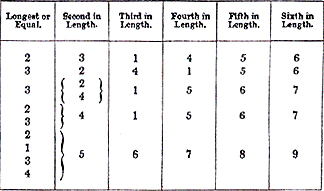
Mr. Romanes urges that there are three cardinal difficulties in the way of natural selection, considered as a theory of the origin of species. These are (1) the fact that all our domestic animals and [[p. 301]] cultivated plants are mutually fertile when crossed, although they often differ in external characters much more than do distinct species; yet natural species, though sometimes differing very little from each other, are nearly always more or less sterile when intercrossed; (2) the swamping effects of free intercrossing upon any individual variation, preventing its ever becoming increased and intensified by natural selection so as to constitute a distinct species; and (3) the inutility of a large proportion of specific distinctions, which consist of small and trivial differences of form and colour, or of meaningless details of structure, which, being of no service to the plants or animals presenting them, cannot have arisen through the agency of natural selection. Mr. Romanes quotes many passages from Darwin's writings admitting the force of these objections, and he shows, more or less successfully, that the explanations Darwin offered are in no case sufficient. Mr. Romanes proceeds to argue that, admitting these objections, natural selection is not, properly speaking, a theory of the origin of species, but that it is a theory of the origin--or rather of the cumulative development--of adaptations. These, he submits, are very different things, because each useful adaptation usually characterises a whole group of species, often a whole genus or a whole family, while the individual species are distinguished from each other, not by adaptive, but usually by trivial, superficial, and altogether useless characters. To account for these facts Darwin and his followers have called in the aid of certain additional causes, such as use and disuse, sexual selection, correlated variability, and, most important of all, the prevention of intercrossing with parent forms. This last cause is brought into action by the isolation of varieties in distinct areas, and its effects are well seen in the distinct but closely allied species that so often characterise oceanic islands. This is thought to prove that, whenever intercrossing is prevented, independent variability is a sufficient cause for the evolution of new species, which will always tend to arise under such conditions, and will be usually distinguished by characters which are not useful to them, and have therefore not been preserved by the agency of natural selection. But, it is argued, such species can never arise without isolation, because intercrossing will continually extinguish all such independent variations of an unuseful kind, and even all such as are useful, unless they occur in considerable numbers together. Except in the case of complete isolation in islands or by great geographical changes in continents, species must have originated in the midst of a parent form, and unless the mutual sterility we find to be a general characteristic of species had appeared at the very beginning to prevent the extinction of all incipient variations by intercrossing, it does not seem possible for these variations ever to have been [[p. 302]] preserved and accumulated so as to form distinct new species. Mr. Darwin's suggested explanation of the whole difficulty is, that a number of similar favourable variations occurring together will afford materials for natural selection to act upon, and will then rapidly increase; while, as to the cause of infertility between the new form and the parent stock, he suggests that varieties occurring under nature will have been exposed during long periods of time to more uniform conditions than have domesticated varieties, and this may well make a wide difference in the result. This view is supported by the opinion of many independent observers, that domestication tends to enhance fertility; while it is a well known fact that in wild species the reproductive system is so delicately balanced that they often become sterile, even with their own kind, when in confinement. Mr. Romanes, however, objects that this suggestion is too vague and too little supported by known facts to explain such a fundamental and almost universal difference as exists between varieties and species in regard to their mutual fertility, and he therefore puts forth his theory of physiological selection. Briefly stated, this theory is, that individual variations in the degree of fertility with the parent form often occur quite independently of any change in external characters. This mode of variation may be either indirect or direct. In the former kind the season of flowering or of pairing may be advanced or retarded, and in either case the individuals so varying can only cross with each other, not with the parent form. In the latter kind the new variety is such that when crossed with the parent form it produces very few offspring, and those offspring are usually sterile; while among themselves these physiological varieties are perfectly fertile as are their offspring. "Once formed as such," he says, "the new natural variety, even though living upon the same area as its parent species, will begin an independent course of history, and, as in the now analogous case of isolated varieties, will tend to increase its morphological distance from the parent form, until it eventually becomes a true species." Mr. Romanes then goes on to argue that, as a rule, these physiological variations are those which occur first, and form the starting-point of new species. He admits that in some cases sterility may be a secondary character, due perhaps to the constitutional change indicated by the external variation; but even in that case physiological selection plays an equally important part, because, if it does not arise, either coincidentally with the ordinary external variation or as a consequence of it, then that variation will not be preserved, but will rapidly be extinguished by intercrossing with the parent type. Having now set forth very briefly, but I believe quite sufficiently and often in its author's own words, what the theory of physiological [[p. 303]] selection is, let us turn back and see how far the facts of variation on which it is founded are adequately and correctly stated; and also endeavour to ascertain with some precision what would happen to the physiological varieties arising independently in the midst of a species, as Mr. Romanes supposes them to do, and whether they could possibly form the usual starting-point of new species. In discussing the "three great obstructions in the road of natural selection," which Mr. Romanes believes to be insuperable by natural selection alone, it will be convenient to take them in the inverse order, leaving the important question of sterility between species to be dealt with after the road has been cleared of the two less important obstructions. (1.) Inutility of Specific Characters.--This forms an essential part of Mr. Romanes' argument as to the necessity for physiological selection to account for the origin of species, but it is only proved to exist by general statements quite unsupported by evidence. He tells us, for example, that an "enormous number" of specific peculiarities are of no use, giving as instances the callosities on the hind legs of horses, or the habit of covering their excrement by some of the cat tribe. In the latter case, however, it is surely not difficult to see a very probable use, for as the excrements in question are exceptionally offensive, their exposure on the surface of the ground might warn such creatures as are preyed upon by them from approaching the haunts of these animals. But this argument from our ignorance is a very bad one when we consider how recently whole groups of specific differences, formerly looked upon as useless, have been brought under the law of utility. The innumerable fantastic diversities in the size, form, colour, and markings of flowers would have been formerly thus classed; but these have now in so great a number of cases been shown to be purposive modifications for aiding in fertilisation, that few naturalists will doubt that all or almost all similar distinctive characters have had a similar origin. So the various kinds of spines and prickles, of hairs or down, of stinging organs or of sticky exudations, once unintelligible, have now been proved serviceable in keeping away "unbidden guests" from the flowers. The life histories of animals in a state of nature have been so much less studied than those of plants that we are quite unable to determine the use of many of the slighter specific characters which distinguish them. But here, too, progress is being made, and many peculiarities can now be shown to be useful which a few years ago would have been classed as of no possible utility to the species. This especially applies to the colours and markings of animals; and having paid much attention to this question I will make a few remarks upon it. It is a very striking fact, the full importance of which has not been appreciated, that almost all animals, when domesticated, produce varieties of colour and markings, often [[p. 304]] exhibiting a great diversity in this respect, whereas the wild species from which they have been derived have each a constant type of colour and marking, and although they not unfrequently produce varieties, such as white or pied swallows, blackbirds, &c., these never increase in numbers as they do under domestication. This implies that the variation is prejudicial to the species, and that the general constancy of colouration we observe in each wild species is a useful character. A long consideration of this subject has convinced me that the usefulness of colour and marking to wild animals arises in many different ways. The most general of all the uses of colour is to serve as a protection to the species from its enemies or to aid in concealing it from its prey; hence the very wide prevalence of protective colouration as instanced, broadly, in the white arctic and sand-coloured desert animals, in the numerous green birds of tropical forests, and, more especially, in the countless insects resembling green or dead leaves, bark, bird's dung, moss, stones, or other natural objects among which they live. The protective character of many of these markings can rarely be understood till the creature is seen in its natural attitude and among its natural surroundings, so that hundreds of species preserved in our museums and cabinets seem to have colours which are altogether unmeaning and useless, owing to our ignorance of their habits and life history. Another kind of colouration was long quite unintelligible, that of creatures which are very conspicuous and often so gaudily coloured as to attract attention; but it is now found that many groups of species thus coloured have a totally different kind of protection in being endowed with such an offensive odour and taste as to be inedible. Whole families of butterflies, moths, beetles, and other insects, are now known by actual experiment to be so protected, and these in every case possess conspicuous colours, or at all events are entirely wanting in those protective hues which characterise most creatures which serve as food to others. Another class of animals possess deadly weapons, like the stings of wasps and the poison fangs of snakes, and these often exhibit conspicuous colours or some other means of warning their enemies that they cannot be attacked with impunity. As illustrations of these forms of useful characters I may mention the glow-worm and fire-flies, which belong to inedible groups, but being nocturnal and soft-bodied would be liable to be seized and injured, if not devoured, without the warning light which tells all insect-eating creatures (after one experience) that they are uneatable. This interpretation of the use of the light was suggested by Mr. Belt and has been adopted by Mr. Darwin. The case of the poisonous snakes is still more curious. Most of these are rather protectively coloured in order that their prey may approach them sufficiently near to be seized, but they are usually characterised by a broad [[p. 305]] triangular head and short tail which sufficiently marks out the tribe of viperine poisonous snakes to reptilivorous birds and mammals. In a few cases, however, they possess a more special warning. The rattle of the rattle-snake and the dilated hood of the Indian and African cobras are of this character, and it is interesting to note that the cobras do not belong to the viper tribe, but have heads and tails of similar form to harmless snakes. In South America there are poisonous snakes of the same family which get protection not by a hood or rattle, but by a style of colouration in alternate rings of black, red, and yellow, quite unlike that of any other snakes in America or in the rest of the world. They are distinguished among other snakes just as the brightly coloured inedible insects are distinguished among their edible allies, and for the same purpose of warning enemies not to attack them. The several cases now referred to cover a great deal of ground, but there remains one of the most important. It may be said, you have shown the use of certain classes or styles of colouration, but these would apply to a great number of species equally well. Why, then, is each species usually different in colouration from all others? The reply to this objection I believe to be, that easy recognition is important to all animals, and especially to those which are gregarious and whose safety largely depends upon their keeping together. My attention was first called to this subject by a remark of Mr. Darwin's, that the principle of protective colouring fails in the rabbit, "for when running to its burrow it is made conspicuous to the sportsman, and no doubt to all beasts of prey, by its upturned white tail." Not believing that any animal could have acquired a character actually hurtful to it without some more than counterbalancing advantages, it occurred to me that, when feeding in the dusk, rabbits run to their burrows on the least alarm, and that it would be very important for those who were furthest off, and especially for the young, to be able to follow the others without any hesitation in a straight line. The upturned white tail thus serves as a useful guide. On looking for other cases of analogous colouring, I was struck by the remarkable fact that a large number of antelopes, which are usually protectively coloured with sandy or earth-coloured tints, are nevertheless rendered conspicuous by large white patches, usually behind or on the flanks, and often accompanied by peculiar white marks on the face, but always different in each species. Mr. Darwin imputes all these markings to the effects of sexual selection, having been first acquired by the males and then transmitted wholly or partially to the other sex. It seems to me, however, much more probable that these markings have been acquired for the purpose of enabling any strayed member of the herd to recognise his fellows, and to be recognised by them. Most of these animals depend for safety on keeping together, [[p. 306]] when they can defend themselves against most beasts of prey; and as each kind will not usually allow animals of another species to join them, it becomes doubly important that every species should have a distinctive marking, especially with desert animals, which are obliged to roam far in search of food and water, and still more when there are many allied species of the same general form inhabiting the same country. It seems not improbable that the many curious differences in the shape, direction, and curvature of the horns of antelopes may have risen from a similar cause, as when these alone were visible they would often serve the purpose of recognition at a great distance. This same idea has occurred to Mrs. Barber, an excellent observer of nature in Cape Colony. She says, "Land birds are for the most part coloured to match the country they inhabit. Some of them, however, possess conspicuous markings, which are of great service to them in their flight, enabling them, if disturbed (especially during the night), to keep together. If, however, they are not in possession of indicative colours (such as white beneath the wings, &c.), they will probably utter some peculiar note or frequent cry, which will answer the same purpose, like that of the fern owl, for instance." This need of easy recognition by each species of its own kind and of the sexes by each other, will probably explain at once those slight diversities of colour and marking, which, more commonly than any other characters distinguish closely allied species from each other, and also the constancy and bilateral symmetry of the colouration of wild animals. For if the same species varied in colour beyond definite limits, and especially if they became piebald or irregularly coloured, great confusion would arise; and it is probable that such irregularities, when they do occur, soon die out, because the normal-coloured individuals refuse to pair with them. I think I have now shown that in a great number of cases the trivial characters that distinguish species from species are, in all probability, useful to them, and may therefore have been increased and fixed by natural selection. This is the more probable if we remember the extraordinarily rigid character of the selection that is always going on among wild animals, from three to ten or a hundred times the minimum population being weeded out every year, so that the very slightest characters, if even at rare intervals affecting the safety of the individual, will be almost sure to be preserved. We must also remember that many slight characters may be the atrophied or rudimentary remains of more important characters which were useful in some ancestral form, but which, being now so very trivial, have not been completely lost by disuse; while sufficient importance has not been given to the constant state of flux and reflux of all organic forms, development and degeneration going on alternately [[p. 307]] and having been many times repeated, so that characters may be partially lost, and then under a change of conditions utilised by a fresh development in a different direction, thus leading to those singular complexities of form and structure, serving purposes which might apparently have been reached in a much simpler and more direct manner. I believe, therefore, that the alleged "inutility of specific characters," claimed by Mr. Romanes as one of the foundations of his new theory, has no other foundation than our extreme ignorance, in the great majority of cases, of the habits and life-histories of the several allied species, the use of whose minute but often numerous differential characters we are therefore unable to comprehend. (2.) Swamping Effects of Intercrossing.--Mr. Darwin's remarks on this subject are as follows:--"Most animals and plants keep to their proper homes, and do not needlessly wander about. We see this with migratory birds, which almost always return to the same spot. Consequently, each newly formed variety would generally be at first local, as seems to be the common rule with varieties in a state of nature; so that similarly modified individuals would soon exist in a small body together, and would often breed together. If the new variety were successful in its battle for life, it would slowly spread from a central district, competing with and conquering the unchanged individuals on the margin of an ever-increasing circle." After quoting this passage, Mr. Romanes objects that a very large assumption is made when the newly formed variety is spoken of as represented by similarly modified individuals--the assumption, namely, "that the same variation occurs simultaneously in a number of individuals inhabiting the same area;" and he adds, "Of course, if this assumption were granted there would be an end of the present difficulty;" and then he goes on to give "reasons" why such simultaneous variations are not likely to occur. But that which Mr. Romanes regards as "a very large assumption" is, I maintain, a very general fact, and, at the present time, one of the best-established facts in natural history. A brief summary of these facts is given in my Island Life (p. 57), and I possess in MSS. a considerable collection of additional facts, showing that simultaneous variation is a general phenomenon among the best-known species of animals and plants. Unfortunately, very few naturalists pay attention to individual variations. They are usually satisfied with describing typical or mean specimens, sometimes noting the amount of variation of size they have met with, but hardly ever taking the trouble to compare and measure scores or hundreds of specimens of the same sex and age, and collected in the same locality, so as to furnish us with direct evidence of the general amount and kind of variation that occurs in nature. One American naturalist, however, has done this; and to [[p. 308]] Mr. J. A. Allen we owe a debt of gratitude for having furnished us, in his Mammals and Winter Birds of Florida, with a complete demonstration of individual and simultaneous variability by a series of minute comparisons and measurements of a large number of common North American birds.1 We have no longer any occasion to reason as to what kind or amount of variation is probable, since we have accurate knowledge of what it is. The following is a brief summary of Mr. Allen's facts. After comparing and measuring from twenty to sixty or more specimens of each of a great number of species, not only as to their general size, but also as regards every external part and organ capable of being measured, he says:--"The facts of the case show that a variation of from 15 to 20 per cent. in general size, and an equal degree of variation in the relative size of different parts, may be ordinarily expected among specimens of the same species and sex, taken at the same locality." He then goes on to show that each part varies to a considerable extent independently of the other parts. The wing and tail, for example, besides varying in length, vary in the proportionate length of each feather, which causes their outline to vary considerably in shape. The bill varies in length, width, depth, and curvature. The tarsus varies in length, as does each toe separately and independently; and all this not to a minute degree, not "infinitesimally," as usually stated, but to an amount that can be easily seen without any measurement, as it averages one-sixth of the whole mean length, and not unfrequently reaches one-fourth. In order to ascertain the amount of independent variability of the different parts, I constructed a series of diagrams from Mr. Allen's tables of measurements, so as to show, by the amount and direction of the curvatures of lines, the variability of each part in a number of specimens of the same species. The comparative lengths of the wing, tail, bill, tarsus, and each of the toes were thus shown for, say, twenty specimens of the same bird; and it was most interesting to note how independent is the variation of each part, so that we may choose either a long wing with a short tail, or the reverse, or both long or both short; a long bill or a short bill with a long leg, or again the reverse; and so with every external character there seems to be no fixed correlation (though a tendency to it is in some cases shown), but each part appears to vary independently of all the rest. As an illustration of this independent variability which can be shown by figures, the following table exhibits the relative lengths of the primary wing-feathers in the myrtle warbler of the United States (Dendrœca coronata), occurring among twenty-five specimens:-- [[p. 309]] Relative Lengths of Primary Wing-Feathers of "Dendrœca Coronata."
Here the figures 1, 2, 3, &c. indicate the wing-feathers numbered in order from the outer one, and we see that either the 2nd, 3rd, 4th, or 5th feather is sometimes the second in length, while the 1st or outer feather is sometimes one of the longest, sometimes third, and sometimes only fourth in length. This is an extreme case, but many other birds give almost similar results. Mr. Allen also gives full details as to the variation of colour and marking, showing that these are not less striking than those of size and proportions; but the most important thing for us in regard to the question we are discussing is the amount of simultaneous variation of the same kind that is constantly occurring. To determine this I formed diagrams, in which each individual was represented by a spot placed on a horizontal line at a point determined by its actual dimensions. It would have been antecedently expected that the great bulk of the spots would be crowded together about a point representing the mean dimensions of the species, but this was by no means the case. Often the central point was not at all crowded with dots, but they were grouped with rough uniformity for a considerable distance on each side of the centre, with a few isolated at greater distances representing the extremes of variation. Hence a species could usually be divided into two portions, with a considerable number of specimens in each showing divergence from the mean condition--the very "simultaneous variation" which Mr. Romanes regards as "a very large assumption." And this result appears more or less prominently whatever characters are compared, so that whether we require modification of wing or tail, of beak, leg, or toes, we always find a considerable number, say from 10 to 20 per cent. of the whole, varying simultaneously, and to a considerable amount, on either side of the mean value. Now, we must remember that these results have been obtained from the comparison of from twenty to sixty specimens only, usually [[p. 310]] collected at one time and place, while nature deals with millions and hundreds of millions of each species, reproduced afresh every few years, with probabilities of variation far beyond those which occur in the very restricted range of one observer. We must also remember that at least 90 to 95 per cent. of the offspring produced each year are weeded out by natural selection (because birds live many years and produce many young each year), so that, during any change of conditions necessitating readjustment to the environment, an ample supply of "simultaneous favourable variations" would occur calculated to bring about that readjustment. And since we have every reason to believe (as I have shown in the preceding section) that the slight specific differences of which these variations are the initial steps are in most cases utilitarian in character, we may feel sure that all useful variations, occurring so frequently, would be preserved and rapidly increased without any danger from the "swamping effects of intercrossing." Having now shown that two of the "great obstructions in the road of natural selection" set forth by Mr. Romanes do not in fact exist at all, we are in a position to consider the effect of the undoubtedly real and important difficulty of the difference between species and varieties in the matter of fertility when intercrossed. (3.) Sterility between Species.--In discussing this question Mr. Romanes assumes that it is almost a universal rule for natural species to be more or less infertile with each other, while domesticated varieties, on the other hand, are almost always perfectly fertile, and sometimes exceptionally so. Supposing this to be a fair statement of the facts, he very naturally objects to Mr. Darwin's explanation of them--that species have been subjected to uniform conditions for long periods--as quite inadequate, urging the great "antecedent improbability, that in all these millions and millions of cases the reproductive system should happen to have been affected in this peculiar way, by the mere negative condition of uniformity;" and further, "that, at the time when a variety is first forming, this condition of prolonged exposure to uniform conditions must necessarily be absent as regards that variety: yet this is just the time when we must suppose that the infertility with the parent form arose." Now let us see whether there is any reason for believing that species which are very closely allied, that is, which have recently been specialised the one from the other or both from a common ancestor, as well as those natural varieties which may be classed as incipient species, agree in being always infertile with each other or in producing infertile offspring. It is important to remark that hybridisers usually experiment with very distinct species, and often with distinct genera, and even such crosses as these not unfrequently [[p. 311]] produce offspring; while in the cases of close allies being quite fertile the conclusion is arrived at that they are really the same species. Dean Herbert's experiments are most instructive in this respect, since they show that in a considerable number of large genera hybrids are perfectly fertile, and not unfrequently more fertile than the parents, while in many cases they produce quite fertile offspring; and he concludes, "that the sterility or fertility of the offspring does not depend upon original diversity of stock; and that, if two species are to be united in a scientific arrangement on account of a fertile issue, the botanist must give up his specific distinctions generally, and entrench himself within genera." He showed that many very distinct species of crinum, hippeastrum, gladiolus, pelargonium, calceolaria, and many other genera were quite fertile when crossed, and often produced offspring which could be propagated indefinitely and have thus formed valuable garden flowers; while other species, more alike externally, either could not be crossed at all, or produced offspring which were sterile; and he thence concludes, "that the fertility of the hybrid or mixed offspring depends more upon the constitutional than the closer botanical affinities of the parents."2 The popular ideas as to the sterility of hybrids are derived from crosses between certain domestic animals by no means closely allied, such as the horse and ass, the canary and goldfinch, or the domestic fowl and the pheasant. To arrive at the common ancestor of either of these pairs we should probably have to go back far into the tertiary period, and trace their diverging progeny through many successive distinct species, so that there is no fair comparison between such crosses and those between domestic varieties, which, however different externally, have all originated within a few thousand years. Really close species which have probably originated by one remove from a common ancestor have never yet been crossed in large numbers and for several generations, under approximately natural conditions, so as to afford any reliable data. The mere fact that not only animals of distinct genera, but even those classed in distinct families--as the pheasant and the black grouse--sometimes produce hybrid offspring in a state of nature, is itself an argument against there being any constant infertility between the most closely allied species, since if that were the case we should expect the infertility to increase steadily with remoteness of descent till when we came to family distinctions absolute sterility should be invariable. I quite agree with Mr. Romanes that on this point experiments are required, and some of those which he has suggested at the conclusion of his paper are well fitted to test the question whether infertility is a cause or a consequence of specific distinction or merely [[p. 312]] a correlative phenomenon. The most direct and easy experiments would be those with plants. We possess a considerable number of native plants which by one school of botanists are classed as species, while by another school they are considered to be only sub-species in process of segregation from a parent form. It would be tolerably easy to determine whether these pairs of allied forms present any definite amount of infertility, which they should do in almost every case to support Mr. Romanes' theory. We have, however, first to consider whether, even if such general infertility exists, it can possibly have been brought about in the way he suggests. The Theory of Physiological Selection.--While fully admitting that variations in fertility are highly probable, and also that there is evidence to show that individual varieties occur which, while infertile with some members of the same species are fertile with others, it yet seems to be quite impossible that such variations should produce the results claimed for them by Mr. Romanes. He says, "If the variation be such that the reproductive system, while showing some degree of sterility with the parent form, continues to be fertile within the limits of the varietal form, in this case the variation would neither be swamped by intercrossing, nor would it die out on account of sterility. On the contrary, the variation would be perpetuated with more certainty than could a variation of any other kind. For in virtue of increased sterility with the parent form, the variation would not be exposed to extinction by intercrossing; while in virtue of continued fertility within the varietal form the variation would perpetuate itself by heredity, just as in the case of variations generally when not reabsorbed by intercrossing." He then goes on to show how, by these means, a species becomes divided into two portions each free to develop independent histories without mutual intercrossing. This statement, with the results deduced from it, sounds feasible when not closely examined; but it really slurs over insuperable difficulties, and when viewed in the light of the known facts of variation and natural selection it will be seen that the supposed results could not follow. Mr. Romanes speaks of this physiological variation as if it were a simple instead of a highly complex form of variation, and as if it might occur sporadically within the limits of a species like some change of colour or modification of form. In order to test this and ascertain what would really happen, we must follow the variety step by step under varied conditions. Let us then suppose that in a large species some one individual is produced that is infertile with the bulk of the species, but fertile with some few individuals of the opposite sex who happen to be what may be termed the physiological complements of the first-named individual. But it will evidently be in the highest degree improbable that these [[p. 313]] complementary pairs should accidentally meet, as, by the hypothesis, there is no external common character distinguishing them from the rest of the species, and if all are sterile with other than their "complements" then all are doomed to almost certain extinction. Now let us suppose that, not one only, but a dozen or a score or even a hundred of such physiological varieties occur at the same time scattered throughout the area occupied by the species, and that each one has some few complementary mates with whom alone it will be fertile. In this case the chances against the right pairs meeting will be almost as great as before, unless we make the assumption that the individuals which vary in the direction of sterility with the bulk of the species all agree in being fertile with any one of the same set of individuals of the opposite sex. This, however, seems to me so highly improbable an assumption that we cannot possibly accept it without direct and cogent proof, since the fact that the different physiological varieties arose in different parts of the area, from distinct parents, and under slightly different conditions, renders it almost certain that each one would require for its complement an individual which would not be the complement of any other. This difficulty is so great that I cannot conceive the possibility of such physiological variations arising sporadically at several distinct points within the area of a species. There is, however, one other way--and it seems to me the only possible way--in which such varieties could arise. The entire offspring of a single pair might, conceivably, be so constituted as to be fertile inter se while sterile with all the rest of the species, and, if they kept together, might form the nucleus of a "physiological variety." But there would evidently be enormous odds against them. For it must be remembered that the weeding-out by the struggle for existence is so terribly severe that only in very rare cases can more than one or two offspring of the same parents arrive at maturity and when this rare event happens it will be essential that they comprise at least one pair of opposite sexes. Then this pair, or pair and a half, after all the chances and changes of early life, after enduring the fierce struggle for existence for several months or for a year, and after each of them has escaped countless perils, and has been driven hither and thither by the need of food, by the inclemency of the seasons, or by the pursuit of enemies, must nevertheless, just at the right time, come together--or become extinct. It must be remembered, too, that there is nothing whatever but chance to bring them together; for there is, by the assumption, no difference of form, or colour, or habit, or instinct, nothing but the one fact--which they themselves cannot possibly know--that unless they happen to meet and pair their particular race will be doomed to extinction. Surely a phenomenon so widespread as the existence of some degree of [[p. 314]] sterility between species cannot possibly have originated in a mode of variation, which, whenever it occurs, is almost certain to die out immediately. I have now shown, by considering carefully the results of the variations suggested by Mr. Romanes, that they could not possibly produce the effects he attributes to them. Yet he has arrived at a diametrically opposite conclusion; for he claims as the special feature of these variations that "they cannot escape the preserving agency of physiological selection. Hence, even if it be granted that the variation which affects the reproductive system in this particular way is a variation of comparatively rare occurrence, still, as it must always be preserved whenever it does occur, its influence in the manufacture of specific types must be cumulative, and, therefore, in the course of geological time probably immense." This most extraordinary statement, which I have just shown to be the very opposite of what would really happen, seems to me to have been reached by ignoring altogether the cardinal fact of the tremendous struggle for existence, and the survival in each generation of only a small percentage of "the fittest." Mr. Romanes' argument almost everywhere tacitly assumes that his "physiological variations" are the fittest, and that they always survive! With such an assumption it would not be difficult to prove any theory of the origin of species. My readers may now reasonably ask whether, having rejected Mr. Romanes' solution of the problem of the general sterility of species as opposed to the equally general fertility of varieties, I have myself any suggestion to make as to how the admitted difficulty may be overcome. I have already stated that some of the more important data for a complete solution are wanting, owing to the very imperfect character of hybridisation experiments from this point of view; but the reconsideration of the whole question to which I have been led by Mr. Romanes' paper (and for which therefore I am much indebted to him) has cleared up some difficulties in my own mind, and has resulted in a provisional explanation which seems to me to be in harmony with most of the facts. This I will now endeavour to explain. Mr. Darwin, in his invaluable work on Animals and Plants under Domestication, has collected a body of curious facts proving a remarkable correlation between physiological peculiarities and colour, both in plants and animals, the bearing of which on this question he appears to have himself overlooked. Dealing first with general physiological correlations, we have the following facts. In Holland red-coloured hyacinths were injured by frost more than plants of any other colour; purple plums are affected by a disease from which green or yellow-fruited kinds are free; in Mauritius white sugar-canes suffer from disease so severely that they have been largely given up [[p. 315]] for red canes, which do not suffer; in France a very fine white onion was found to be especially liable to fungus; in Malaga green grapes had the vine disease severely, while red and black sorts did not suffer at all. Analogous facts in animals are that white terriers suffer most from distemper; white or white-spotted horses are poisoned by eating mildewed vetches, which did not injure brown or black horses; in the Tarentino black sheep are kept because white sheep are poisoned by eating the Hypericum crispum which abounds there; in Virginia black pigs alone are kept, because they alone are not injured by the poisonous paint-root; white chickens are found to be most subject to the gapes; while in France the yellow-cocooned silkworms have fungus disease much more than the white-cocooned varieties.3 Here we have a very remarkable series of cases showing that the whole constitution of animals and plants is often profoundly modified in correlation with changes of colour, while no such constitutional changes have been observed to accompany such modifications of form and structure as are usually met with in varieties or allied species. We are taught by these facts that colour is an important character, physiologically; and as we know it to be so frequently modified for protective or other utilitarian purposes, we can see what a powerful selective agency it may become, especially as we may be sure that numbers of less obtrusive correlations than those which seriously affect health and life must have remained unnoticed. But in the same work Mr. Darwin furnishes us with another set of correlations, in which infertility or complete sterility is directly correlated with diversity of colour. The red and the yellow varieties of maize were found by Gärtner to be almost completely sterile when crossed; the yellow and the white varieties of mullein will not cross, although many distinct species, if both yellow or both white, are perfectly fertile when crossed; the differently coloured varieties of the hollyhock are raised by nurserymen in rows close together and never hybridize, each sort keeping distinct, although they are visited by bees; and, lastly, the blue and the red pimpernels, considered by most botanists to be the same species since they present no differences of form or structure, are yet completely sterile when crossed.4 Among animals no experiments have been made to show how colour affects the sterility of crosses, but there is ample evidence that the same result is brought about by the disinclination of differently coloured races to pair together. In Paraguay and in Circassia it has been noticed that feral horses of the same colour and size usually breed together; in the Faröe Islands the black and [[p. 316]] the white sheep keep in separate flocks; in the Forest of Dean and in the New Forest dark and pale herds of deer do not mingle together; while pigeon-fanciers agree that if pigeons were allowed freedom of choice they would pair with their own sort exclusively.5 Many of the facts here summarised rest upon the testimony of more than one good observer, while in several cases they were confirmed by Mr. Darwin's own observations; and they certainly demonstrate the great importance of colour, both as a physiological selective agency in certain localities, and as correlated with varied constitutional differences, with disinclination to pair together in animals, and with actual mutual sterility in plants. But it is a matter of common knowledge to naturalists that differences of colour or markings form the very commonest of the distinctive characters between closely allied species, while they also frequently characterise the varieties of the same species. From a utilitarian standpoint colour is, as I have shown, one of the most important of specific characters, serving in infinitely varied ways the several purposes of concealment, of warning, and of recognition; and, therefore, a difference of colour is almost sure to arise whenever, by natural selection, a species is becoming adapted to any change in its environment. Now taking into consideration the remarkable facts above enumerated, it is surely a not improbable supposition that change of colour is usually accompanied by some amount of sterility, and of disinclination to pair in the case of animals; and that it thus furnishes the required starting-point of that physiological distinction which becomes more marked when, by successive variations and adaptations, the original varieties of one parent form have become changed into distinct and well-marked species. The extreme generality of colour as a specific distinction, is in perfect accord with the generality of some amount of sterility between distinct species; and we thus have a vera causa co-extensive with the effect produced. In conclusion, I do not deny that varieties which exhibit no other distinctive character than sterility with the bulk of the parent species may arise, but I claim to have shown that such varieties are at an immense disadvantage, and could hardly by any possibility be preserved and increased till they were required to form the nucleus of a new species. On the other hand, I have shown that sterility or infertility is actually, in many cases, correlated with colour-variations, while this very character of colour-variation is the most frequent mark of closely-allied species or sub-species. It is, therefore, by means of a study of this class of facts that I believe the true solution of the problem of the sterility of hybrids will be discovered.
1This
work had not reached England when the last edition of The Origin of
Species was passing through the press, but it is referred to in the
Descent of Man. [[on p. 308]]
The subject of Wallace's attack, Romanes' 1886 article in the Journal of the Linnean Society, is presented at: http://post.queensu.ca/~forsdyke/romanes1.htm. Although at that time Romanes' health was excellent, he had only eight years to live. He died a few days after his forty-sixth birthday of what seems to have been a brain tumour. In contrast, Wallace, then sixty-three, was to live to 1913. The Linnean Society lecture was the first salvo in an eight year public "debate" between Romanes and the evolution establishment, which was no less colourful than its equivalent a century later between Stephen Jay Gould and the modern evolution establishment. The issues were basically the same, with Romanes (1886) and Gould (e.g., 'Is a New and General Theory of Evolution Emerging?', Paleobiology 6: 119-130, 1980) both advocating a higher level of selection involving agencies other than the natural selection of Darwin and Wallace. Remarkably, Gould himself contracted cancer at about the same age as Romanes, but with modern therapies survived another two decades to die in 2002 on 20 May (which happened to be Romanes' birthday). Of necessity, Romanes and Gould could only describe their agencies in abstract terms (see Gould, The Structure of Evolutionary Thought, 2002) . Romanes, a physiologist by training, called his "physiological selection." New bioinformatic analyses of the DNA sequence data that emerged from various genome projects in the 1990s suggest, however, that we may now be able to flesh out these abstractions in chemical terms (see Forsdyke, The Origin of Species, Revisited, 2001). |