
http://people.wku.edu/charles.smith/biogeog/MATT1913.htm
Certain Theoretical Considerations Affecting
Phylogeny and Correlation1
by W. D. Matthew (1913)
Editor Charles H. Smith's Note: Original pagination indicated within double brackets. Notes are
numbered sequentially and grouped at the end, with the page(s) they originally appeared at the
bottom of given within double brackets. Citation: Bulletin of the Geological Society of America
24 (1913): 283-292.
[[p. 283]] (Read before
the Paleontological Society December 30, 1912)
CONTENTS
Theory of centers of dispersal . . . . . . . . . . (page)
283
Admixture of blood through migrations . . . . . . . . .
. 284
The English race as an illustration . . . . . . . . . . 284
Applicability of the principle to animals . . . . . . . . . . 285
Effect of interrupted migrations . . . . . . . . . . 287
Effect of remoteness from dispersal center . . . . . . . . . . 287
True nature of a phyletic series . . . . . . . . . . 288
Correlation of homotaxial stages of a phylum . . . . . . . . . . 288
Correlation of Cenozoic mammal faunæ . . . . . . . . . . 290
The Pampean fauna: Equidæ and ground sloths . . . . . . . . . . 291
THEORY OF CENTERS OF DISPERSAL
In a former communication to the Society2 I discussed the evidence for the continuity of evolution from the standpoint of vertebrate paleontology. The theory adopted to interpret the evidence was that of evolution of each race in a center of dispersal, and spread from there in successive waves of migration to other regions. The present contribution aims to discuss certain phases of this theory in their effect on the structure and affinities of a phyletic series in a region remote from the center of dispersal.
The considerations to be presented and their effects on the composition and affinities of a race are familiar to modern ethnologists. They are recognized facts of the dispersal of the human races, and their effect on the affinities of each race is clearly seen and universally recognized. [[p. 284]] Their application to zoological, and especially to paleontological, studies is, I believe, no less exact, but has received little attention.
ADMIXTURE OF BLOOD THROUGH MIGRATIONS
In presenting the theory of dispersal in its application to fossil vertebrates, it is convenient to speak of a series of waves of migration proceeding outward from a center of dispersal. In fact this is a somewhat misleading phrase. What must really have occurred is rather a more or less continuous stream of new blood pouring into the region considered, mixing with the older blood and gradually displacing it. If the course of migration be uninterrupted and the environmental pressure be more or less continuous in its action and direction, the progress of a race in a given region will be due partly to the infusion of new blood from the adjoining region nearer the center of dispersal, partly to the adaptive influence of the local environment, partly to the substitution of new species or genera arriving from regions nearer the center of dispersal and which either were originally or have become infertile with the autochthonous species.
In discussion of the evolution of a phylum, the second and fourth of these influences have usually been considered. But it would seem that all four have frequently, if not generally, played an important part in determining the evolutionary progress of a phylum in any locality. The representatives of a phylum at any one stage in its development in a given region do not consist of one or several homogeneous species or pure races, but of series ranging from archaic autochthones to recent immigrants, more or less broken into nodes or separated groups by local segregation or the partial extinction of intermediate members of the series.
THE ENGLISH RACE AS AN ILLUSTRATION
Let us consider, first, what we know to have been occurring in the migrations of the human race and its effect on the population of Great Britain. When this island first appears in history its population is somewhat heterogeneous. The inhabitants of the south and east are closely allied to the Gauls. Northward are traces of Scandinavian and in Ireland of Iberian affinities. The Roman occupation, while it checks migration for a time, does not introduce any important new ethnic element, but toward its close the normal course of migration is resumed and an ever increasing flood of Platt-Deutsch peoples pours in. The effect of the hindrance of migratory movements during Roman times is seen in a partial disinclination to free admixture, and a certain degree of segregation [[p. 285]] between the Teutonic invaders and Celtic aborigines is evident, and indeed persists to some extent today. Following the Saxons and Angles came Jutes and subsequently Danes, forming the dominant element of the eastern coast, while the Norman-French spread a new layer of population over the whole country. Subsequently the south and east receive new increments of population from the Netherlands and later from France. There is likewise a considerable movement of population from Scotland south into England and the north of Ireland, from England into Ireland, and more recently from Ireland back into England. Setting aside the concentration into the cities and considering only the rural population, these, with other minor movements, afford the necessary data for interpretation of the wide variations in physical type in different parts of the British Isles. The unity of type among Englishmen is based on language, customs, and conventions, in some degree perhaps on natural environment; but it covers a wide diversity of blood. The Englishman is not merely of mixed race, but the admixture varies greatly locally. This is almost equally true of every civilized race, and it is to a very large extent true of every uncivilized race.3
The evolution of the human race in any one region has been chiefly through successive infusions of new blood, not through the replacement of one race by another; and this is true of barbaric races. The dominant race absorbs the remnants of the older blood or is absorbed by autochthones, but the continued renewal of new invading elements disturbs the homogeneity of the population before it is complete, and enables a very archaic strain to be preserved to some extent in association with new elements, which would not have fused with the ancient blood except through the intermediary of successive previous invasions of intermediate stocks.
APPLICABILITY OF THE PRINCIPLE TO ANIMALS
The evolution of mammalian races must have proceeded, so far as I can see, on much the same lines. If so, when we come to make detailed studies of the evolution of mammalian phyla, these controlling circumstances must be carefully kept in mind. If these principles be true, then in the normal course of evolution of a race in a given region, each stage [[p. 286]] will consist of a complex of groups of varying blood-mixture, inextricably tangled up, each individual being a product of indirect hybridism of many ancestral strains, some of which would be generally regarded as distinct species.
The old Linnæan species concept was founded on supposed impossibility of blood relationship between mutually infertile forms; the current concept makes the specific group dependent rather on local or preferential segregation than on infertility for their distinctness. But if the migration and dispersal of a race involves this sort of indirect hybridism, even the broader concept of the Linnæan species will not secure complete racial distinctness to the subphyla of a polyphyletic group. The horse and the ass are today immiscible, yet the wild horses, if we assume that they are a later invasion, occupying territory formerly peopled by asses, probably have ass blood in their veins, and individuals should present varying proportions of characters due to that inheritance. It is not indeed the characters of the modern ass that should appear, but the characters of (1) the ass stage in the center of dispersal of the Equidæ, more remote and more modified by evolutionary change, but reinforced by (2) admixture of the remnants of the former equine population of the region invaded by the true horses.
The net results of this process of admixture will be to produce a reactionary effect on the characteristics of an invading group as it reaches regions remote from its center of dispersal, through continual and progressive absorption of its predecessors in the dispersal of the race. This will be added to the effect of the local environment and of earlier environments which it has passed through; and all these factors dependent on shifting habitat must be added to those which operate in a fixed habitat to produce or control evolutionary progress in a race of animals evolving and spreading out from a center of dispersal.
Whether the characteristics be inherited according to Mendelian laws, or be fluctuating variations tending to revert to type, will not affect the results of this continuous admixture and readmixture, so far as I can see. Nor will it make any particular practical difference from the present point of view whether Mendelian characters arise suddenly or gradually. Under the stated circumstances, it will not be possible to eliminate a Mendelian character any more easily than a non-Mendelian character. The problem is far too complex and the data are too scanty to afford conclusive evidence as to such niceties. We can obtain and be sure of approximate results and these only. Thus the three strains which Ewart has shown to exist in the domestic horse present a convenient approximate statement of the facts; but the exact sources are probably vastly [[p. 287]] more complex, the original strains are themselves composites of complex origin, and the present habitat of the nearest living representatives of these strains is not their original home.
EFFECT OF INTERRUPTED MIGRATIONS
The problem has thus far been treated on the assumption of free opportunity for dispersal and continuous environmental pressure, producing a continuous stream of migration. But in fact we must expect periodic interruption of greater or less duration in this movement, due either to barriers interposed or to temporary cessation or reversal of the environmental pressure, of whatever nature this may be. These interruptions, if short, will produce nodes; if longer, actual breaks in the stream as it moves forward in time and space. The nodes in the further progress of the stream will tend to disappear; the actual breaks involving infertility may, if sufficiently great, be permanent. In a region far from the center of dispersal the arriving stream will preserve completely or partially all the nodes and breaks due to interruptions at former stages of its progress, the nodes tending to disappearance, the well marked breaks to sharper accentuation. Hence the farther away we are from the center of dispersal the more the race will be broken up into sharply differentiated species or genera, and the less in proportion will there be of nodes or imperfectly differentiated and anastomosing groups.
EFFECT OF REMOTENESS FROM DISPERSAL CENTER
Furthermore, a race remote from its center of dispersal will have passed through many different local environments on its way, and each of these, especially during interruptions of the migration movement, will have tended to impress on its members a local adaptive radiation, which will increase and intensify the diversity of the resultant in its further course. The influence of the typical evolution of the race proceeding in its center of dispersal will be more remote; that of the various divergent adaptations passed through will be more dominant, as the race is more remote from its original home.
Near the center of dispersal, therefore, we should expect to find the specific and generic groups of a family difficult to distinguish, intergrading a great deal and held more strictly to type. Far from the center of dispersal we should expect more sharply differentiated species or genera, more diversity of type, and more aberrant forms.
[[p. 288]] TRUE NATURE OF A PHYLETIC SERIES
In view of the above considerations, what is the real nature of a phyletic series, such as may be found in the successive geologic strata of any region? Evidently its successive stages are not wholly in genetic sequence unless we are in the exact center of dispersal of the group. Nor are they wholly successive immigrants arriving unaltered from that center. Each stage is a complex and non-homogeneous group derived from earlier and later strains of immigrants, modified to varying degrees by the various environments through which they have passed. To disentangle such a complex in any accurate and exact way is humanly impossible, even where our material is abundant. Their approximate relations may be presented in the form of a polyphyletic succession, but all such subphyla must needs be more or less arbitrary and inexact. It is only in the larger phyla that the arrangement accords with the real genetic sequence.
CORRELATION OF HOMOTAXIAL STAGES OF A PHYLUM
There is one phase of this discussion that bears on the question of correlation. It is customary to regard the appearance of equivalent stages in a phylum in the formations of widely separated regions as the most conclusive evidence of synchronism. This may be true where the regions are equidistant from the center of dispersal of the group in question. Where they are not, two considerations affect the conclusion. First, there is the distance in time of migration of the race from the dispersal center to the two regions compared, and this may be considerable, even in a geologic sense. Second, there is the reactionary influence of previous stages in the evolution of the group absorbed into the advancing new stage on its way. This latter is more considerable in its influence than might at first be supposed on species remote from their place of origin of the group. By assuming certain special cases, it is possible to calculate what effect it has.
In the accompanying diagram the horizontal distance left to right represents geographic migration from the center of dispersal. The vertical distance from below upward represents geologic time-relations. A, B, C, D, E, F represent successive geographic varieties or species, formed by the influence of the local environments when the dispersal of the race begins. 1, 2, 3, 4, 5, 6, etcetera, represent successive geological mutations or phyletic stages in the center of dispersal. The upward trend of the line A-F covers the time required for a migrating stage to reach the [[p. 289]] region F from the center of dispersal. The line of direct genetic sequence at the center of dispersal will be represented by A, Al, A2, A3, etcetera.
According to the theories set forth above, we will have at region B first the descendants of A, becoming modified by the B environment; at region C the descendants of A, partly modified by B and C environments, and so on. The next mutation in region B will consist of the incoming
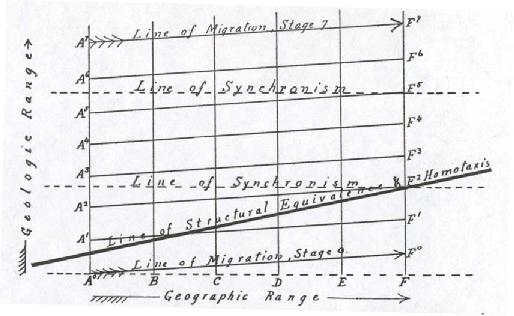
Figure 1.--Relations of homotaxial and synchronous Stages in a widely dispersed Phylum
migrants of the A1 stage, mixing with and partially displacing the original B, derived from A. Assuming as a special case that the newcomers displace three-fourths of the earlier stage and absorb the remaining fourth, then the B1 mutants will contain (as an average) 3/4 of A1 blood, 1/4 of A blood already somewhat modified by B environment, or B1 = 3/4 A1, 1/4 A.
Carrying on the same special case, we will have
C1 = 9/16 A1 + 3/16 A + 1/4 AB.
The modifying influence of the B environment is really somewhat greater than this.
At F1 the approximate figures will show
1/4 A1 influence (23.73 per cent).
1/6 A blood.
The remainder is A blood modified by the environments B, C, D, and E.
[[p. 290]] In other words, the successive remnants of the earlier stage absorbed by mutation 1 on its journey from A to F have brought it nearly back to the earlier phyletic stage, although the new stage displaced most of the individuals of the earlier stage in each region it traversed.
If now we calculate the affinities of the next phyletic stage in its progress from A2 to F2, we will find that
B2 = 3/4 A2 + 3/16 A1 + 1/16 A,
and when we arrive at F2 we find that the approximate percentages are
A2 = 24 per cent.
A1 = 29 per cent.
A = 13 per cent.
AB = 12 per cent.
ABC = 10 per cent. A and modifications, 49 per cent.
ABCD = 8 per cent.
ABCDE = 6 per cent.
In terms of structure this means that the representatives of the third phyletic stage by the time they reach region F will have reverted so far as to be less advanced than the second stage, and also undergone a large amount (36 per cent) of atypical modification from the intermediate environments.
If we were correlating the fauna of region A with the fauna of region F on the usually accepted basis of identical or equivalent species, we should conclude that the appearance of F2, which is between A and A1 in its structural progressiveness, should be correlated with strata in region A between A and A1. In point of fact it is later than A2 by the amount of time required for a migrating stage to traverse the distance A to F.
CORRELATION OF CENOZOIC MAMMAL FAUNÆ
These considerations have some bearing on the correlation of Tertiary horizons in Europe and the United States; but so much of our mammalian fauna is derived from dispersal centers more or less equidistant from these two regions that no serious error can ensue from neglecting them. The case is somewhat different when we correlate the faunæ of the southern continents with those of the Holarctic realm. In these we should evidently make some allowance for the time required to reach regions remote from the dispersal center and for the reactionary effect of absorption of more primitive stages. Both will tend to make southern [[p. 291]] representatives of northern groups seem more ancient than they really are. Conversely, northern representatives of southern groups will be more primitive than their contemporaries in the south, and in certain correlations the comparison of the two may furnish the mean homotaxial relation representing actual synchronism.
THE PAMPEAN FAUNA: EQUIDÆ AND GROUND SLOTHS
Equus, for instance, appears first in the Upper Pliocene in the Old World, Lower Pleistocene in North America, Middle Pampean in Argentina. It is of Holarctic, probably Palearctic origin. In South America it is accompanied and partly preceded by peculiarly modified and somewhat less progressive genera, Hippidion and Onohippidion, in which influence of North America predecessors is apparent; but the modification seems to be partly due to South American environment as well. We would conclude that the Middle Pampean is later than the base of the Pleistocene, but how much later would be difficult to say.
On the other hand, the Ground Sloths are of South American origin. The genus Mylodon appears in the Monte Hermoso beds, below the Pampean in South America, in North America in the Lower Pleistocene, and is accompanied here by Megalonyx, less progressive but modified peculiarly, appearing somewhat earlier, and bearing almost the same relations to Mylodon that Hippidion does to Equus. Megalonyx also appears earlier in North America (Blanco or Middle Pliocene) and is unknown in South America, although evidently descended from the Miocene Megalonychidæ, just as Hippidion is from the Miocene Pliohippus of North America. From this we would conclude that the Monte Hermoso beds underlying the Pampean are older than our Lower Pleistocene. By comparing this result with that derived from the Equidæ the equivalence of the Pampean with the Pleistocene seems assured.
A similar method of treatment of the various groups common to the Pampean and Pleistocene fauna of South America and Holarctica yields confirmatory results. These are:
Canidæ, Ursidæ, Felidæ, Procyonidæ, Cervidæ, Camelidæ, Equidæ, Tapiridæ, Mastodontidæ, Muridæ, etcetera, of Holarctic origin.
Erethizontidæ, Mylodontidæ, Megalonychidæ, Glyptodontidæ, Tatusiidæ, etcetera, of Neotropical origin.
It is not often that so fair an opportunity as this is afforded for correlation in far distant regions. More commonly the isolation of the region has prevented or impeded faunal interchange; the migration has [[p. 292]] not been faunal, but more or less sporadic, and all in one direction; and local adaptive radiation has caused progressive evolution, the resulting parallelism and convergence adding new difficulties to the correlation problem. It is doubtful whether such correlations can ever be correctly and accurately made except on diastrophic criteria, and for this method our data are not yet sufficient.
_________________________
Notes Appearing in the Original Work
1. Manuscript received by the Secretary of the Paleontological Society January 28, 1913. [[on p. 283]]
2. Continuity of development. Popular Science Monthly, Nov., 1910. [[on p. 283]]
3. The heterogeneous and complex composition of all human races is strongly emphasized in Boa's recent book, "The Mind of Primitive Man." Dr. Boaz compares it with the composite character of domesticated species of animals, and contrasts it with the supposed purity of wild races. But I do not think that there is any such contrast. Wild animals, like domestic, are of mixed and composite blood, but their admixture has gone on under natural law and conditions, producing certain limitations and uniformities of the resultant that distinguish it from the artificial admixtures present in domesticated races. [[on p. 285]]
*
*
*
*
*