
http://people.wku.edu/charles.smith/biogeog/GRIN1914.htm
The Colorado River as a Highway
of Dispersal
and Center of Differentiation of Species / The Colorado
River
as a Hindrance to the Dispersal of Species /
The Problem of Barriers with Regard
to
Birds and Mammals
by Joseph Grinnell (1914)
Editor Charles H. Smith's Note: The middle three sections
from the monographic study An Account of the Mammals and Birds of the
Lower Colorado Valley, with Especial Reference to the Distributional Problems
Presented. Original pagination indicated within double brackets. Citation:
University of California Publications in Zoology 12(4) (1914):
97-110.
[[p. 97]] THE COLORADO RIVER AS A HIGHWAY
OF DISPERSAL
AND CENTER OF DIFFERENTIATION OF SPECIES
According to Gilbert and Scofield (1898, pp. 487, 488) the peculiarities of the fish fauna of the Colorado River bespeak a very long period of absolute isolation. A remarkably high percentage of its fishes are specifically distinct from those of the other river basins of western North America. The same is to be said of the riparian birds and mammals.
So far as known to the present writer, none of the species listed with a star in the following table ranges beyond the confines of the Colorado River basin, including of course its various tributaries and distributaries such as the Gila and New rivers, except sporadically, or as accounted for by distal invasion through passes or along sea-coastal tracts. In other words, the Colorado River has been in existence so long that the conditions imposed by its presence have figured in the differentiation of representative species of several families, both mammalian and avian.
The great age of the Colorado River is indicated geologically by the vast extent and slow rate of the erosion involved in the formation of the Grand Cañon. This time-element is justly inferred to have been an essential condition in the formation of these species.
SPECIES WHICH BELONG TO THE RIPARIAN BELT,
AND WHICH ARE
THEREFORE HEMMED IN BY THE PARALLELING DESERT TRACTS.
STARRED SPECIES ARE PECULIAR TO THE COLORADO SUBFAUNA
BREEDING BIRDS
Agelaius phoeniceus sonoriensis*
Melospiza melodia saltonis*
Pipilo aberti*
Guiraca caerulea lazula
Piranga rubra cooperi
Vireo belli arizonae*
Vermivora luciae*
Dendroica aestiva sonorana*
Toxostoma crissale*
MAMMALS
Castor canadensis frondator*
Peromyscus maniculatus sonoriensis
Sigmodon hispidus eremicus*
Reithrodontomys megalotis deserti
Neotoma albigula venusta*
Ondatra zibethica pallida*
Mephitis estor
Procyon pallidus*
It may not be amiss to consider these riparian species somewhat with regard to origin. By confining our attention to the north-and-south valley of the lower Colorado, this becomes, with a knowledge of the general status in North America of each group represented, a [[p. 98]] comparatively simple matter. The axiom holds, that, because of the ever-shifting location geographically of associational, faunal and zonal conditions, every single element or line of descent, now represented in the biota of any one locality must have come either in its present form or in some antecedent one from somewhere else. This is certainly true of all terrestrial life. Elevation and depression have worked like a seesaw in dislocating faunas. The Colorado valley is a trough, hemmed in associationally on either side, and only capable of influx of riparian elements at either end. Therefore the riparian species of the Colorado fauna can have entered the area under consideration from only two directions: from the north and from the south.
Only two species are clearly seen to have entered the Colorado valley from the north: Castor canadensis frondator, and Ondatra zibethica pallida. The following species or subspecies are believed to have come in from the south: Pipilo aberti, Guiraca caerulea lazula, Piranga rubra cooperi, Vireo belli arizonae, Vermivora luciae, Toxostoma crissale, Sigmodon hispidus eremicus, Neotoma albigula venusta.
In the remaining seven cases (Agelaius, Melospiza, Dendroica, Peromyscus, Reithrodontomys, Mephitis and Procyon) no grounds are apparent to the writer for assigning either one over the other direction of invasion, and this in spite of whatever may be the marked Austral or Boreal distributional affinities of each group concerned. The tide of invasion may in fact in these species have tended in one direction at one period, in the opposite at another; or, as in Peromyscus maniculatus sonoriensis, the Colorado valley may have acted continuously as a narrow bridge where have met and mingled descent-lines from both the north and the south.
The obvious fact that southern representatives prevail over northern ones is clearly attributable to the present zonal condition obtaining in the region, namely Austral, in its Lower Sonoran division. And evidence elsewhere assembled (Grinnell and Swarth, 1913, p. 383) points towards an increasing temperature throughout the region. This would result in decreasing the favorableness to Boreal forms and increasing the availability of the region for immigration of Austral types. Consideration of the xerophilous vertebrates as well as of the riparian ones leaves little doubt in the writer's mind but that this has been the actual course of events. The northern contingent is on the wane, the southern in the ascendency.
The query presents itself: is the Colorado fauna full? Are all the ecological niches, which are available in this area and which have [[p. 99]] occupants in other regions, occupied here? Probably not, for the intervention of barriers has doubtless prevented the invasion of types which, if they could have once gotten there, would have thriven and assumed a place as endemic elements in the fauna. Sporadic incursion, as of migrants among birds, and strays among both birds and mammals, do not appear to the writer to figure in such a process. Rather must it be a progressive invasion of the species en masse, acquiring, it may be, adaptive modifications as it proceeds. In other words, the conquering of the land is the combined result of the facilities offered by it plus the relative amenability of each species concerned.
The twelve riparian species and subspecies peculiar to the Colorado fauna vary much in degree of difference from their near relatives which occupy adjacent differentiation areas. These varying degrees of difference might be interpreted as measures of the periods of time elapsed since the entrance into the region of each of the types involved. That this conclusion is poorly grounded is evident upon consideration of the various other elements which must figure in the process of species formation. Among these may be suggested: degree of isolation, divergence of homologous associational conditions in the new region from those in the ancestral, and inherent susceptibility to adaptive modification in each of the species concerned.
In the problem of the origin of the riparian portion of the Colorado fauna we seem to have to do with an accentuated kind of isolation. For, as already asserted, there is such a thing as more and less isolation. In the region here considered, possessing extreme associational contrast, we find the ordinary geographic, or more properly speaking, physiographic, isolation coupled with associational isolation. In consequence of this extra favorable contingency, differentiation of species may have progressed with particular celerity, with such distinct forms to show for it as Pipilo aberti, Vermivora luciae, Toxostoma crissale, and Procyon pallidus.
The axiom has presented itself in this connection that the more restricted a species is associationally, that is, the more confined to a narrow range of associational conditions, the more subject it is to the important factor of isolation; hence the more liable to give rise to new incipient strains in different parts of its general range.
An assertion which seems at first glance opposed to the above is: that the less restricted a species is associationally, that is, the more widely adaptable to varying conditions, the more numerous the chances [[p. 100]] for local operation of isolation, because more opportunity for radial dispersion to carry the species into distant localities and under extreme conditions, and for the ultimate interposition of more or less efficient barriers. The factor of distance might here replace the operation of associational restriction in segregating descent-line plexuses.
These are apparently incongruous notions, but the following conception tends to harmonize them; namely, that, granting the three totally different orders of distributionally limiting factors (zonal, faunal and associational), it is probable that different species are restricted unevenly with respect to the three; thus a certain wood-rat (Neotoma intermedia desertorum) is restricted faunally, but ranges widely through zones and associations; a certain wren (Telmatodytes palustris and subspecies) is tightly restricted associationally, but ranges widely through faunas and zones; a xerophilous genus of rodents (Perognathus) is closely restricted zonally and associationally, but ranges rather widely as to fauna. So that both the above assertions might well be true of a single animal historically and even, in different parts of its range, simultaneously! Certainly the first serves in explanation of the multiplicity of geographic races or species in several widespread groups of birds and mammals.
THE COLORADO RIVER AS A HINDRANCE TO THE
DISPERSAL OF SPECIES
Bats and most birds find in the Colorado River no hindrance whatever to individual travel. Freedom of aerial locomotion gives them superiority over any obstruction on the general level of the country they inhabit. It is possible that in a few of the resident birds of limited flight individuals do not regularly cross the main stream, though they readily could do so if such an exigency as that of fire sweeping the bottom lands should drive them to it. Geococcyx californianus, Pipilo aberti and Toxostoma crissale are birds which probably do not often cross the river under normal circumstances.
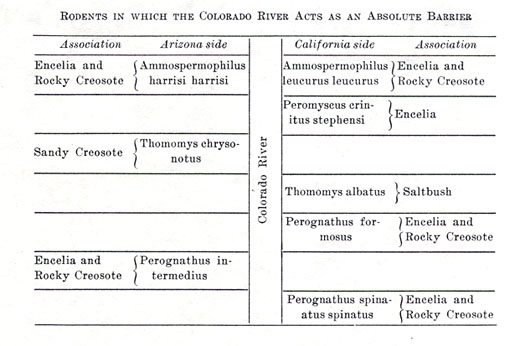
Among mammals, carnivores are usually of much wider foraging range than rodents. From all the data available it appears that none of the carnivores, not even the cats, are averse to swimming the river if need be. Among rodents, however, our work showed a number of cases in which the Colorado River had effectively checked the distribution of species. The following tables show the situation as regards all the rodents of the region (see also figs. A, B).
[[p. 101]]
RODENTS WHICH OCCUR ON BOTH SIDES OF THE COLORADO RIVER, BUT WHICH SHOW
SLIGHT, ALMOST IMPALPABLE, DIFFERENCES ON THE TWO SIDESSpecies . . . . . Association
Citellus tereticaudus tereticaudus . . . . . Sandy Creosote and Saltbush
Dipodomys deserti deserti . . . . . Sandy Creosote and Saltbush
Perognathus penicillatus penicillatus . . . . . Saltbush and Sandy Creosote
RODENTS WHICH ARE APPARENTLY IDENTICAL ON THE TWO SIDES OF THE COLORADO RIVER Species . . . . . Association
Castor canadensis frondator . . . . . River
Peromyscus maniculatus sonoriensis . . . . . All Riparian
Peromyscus eremicus eremicus . . . . . Saltbush and Sandy Creosote
Sigmodon hispidus eremicus . . . . . Willow and Tule
Reithrodontomys megalotis deserti . . . . . Tule and Willow
Neotoma albigula venusta . . . . . Mesquite and other Riparian
Neotoma intermedia desertorum . . . . . Encelia
Ondatra zibethica pallida . . . . . River
Dipodomys merriami merriami . . . . . Saltbush and Sandy Creosote
Perognathus bombycinus . . . . . Sandy Creosote
Lepus californicus deserticola . . . . . Creosote and Saltbush
Sylvilagus auduboni arizonae . . . . . Quailbrush and Mesquite
[[p. 102]] The fact
is apparent that only members of the strictly desert associations are
stopped at the river. And of these the species of the most remote
associational position are, with one exception, Neotoma intermedia
desertorum, most effectively delimited. Also degree of isolation
is in a measure commensurate with amount of difference between
forms of the same genus.
It is pertinent to inquire how the Colorado River acts as a barrier to those species affected. It appears that in every one of the eleven cases the animal in question has no need to visit any water-supply. All are species capable of maintaining successful existence without a drop of water other than that obtained by chemical elaboration from their food. In our three months' experience we did not once find evidence that any individual of any of the eleven species in question had visited the river's edge.
Furthermore, to the best of our knowledge, all the species are of limited foraging range. In the case of the two diurnal chipmunks, Ammospermophilus harrisi harrisi and Ammospermophilus leucurus leucurus, which could be seen, it was seldom that an individual was come upon more than fifty yards from its burrow. In the case of Perognathus, which carefully closes the mouths of its burrows for the day, after its night's activity abroad, it was impossible to secure definite information on this score except as afforded by trapping; but the writer's impression is that it, too, does not ordinarily venture many rods from its retreat. Individuals doubtless travel farther at times of rutting, but it is likely that even then the limits of the native association would not be far transgressed.
It is further to be noted that those species finding an insuperable check at the river are all closely confined to one general kind of associational environment, even though two minor associations, as here defined, be occupied. The river plus intervening associations of an unfavorable nature constitutes the total barrier to the rodents in question.
It is true that the element of distance here implied is reduced to a negligible quantity where hills closely abut upon the river channel. But the major part of the river's course, probably four-fifths of it below the lower end of the Grand Cañon, is through valleys of varying width, occupied by riparian associations most adverse in essential ecological particulars to the species of the upland deserts adjacent.
Along the remaining fifth of the river's course, where the banks rise abruptly and are continuous with the adjacent hill slopes, with [[p. 103]] either no trace of riparian tracts or only narrow or interrupted representations, it would seem that chance is afforded for such mammals as the desert chipmunks and pocket mice to encounter the river itself, with all intervening factors removed.
At our base camp (no. 4) at The Needles, our measurements showed the actual width of the stream to be 450 feet at the rather low stage of water obtaining at that time (March 4, 1910). At high water the river could not have been much more than 150 feet wider (see sectional profile, fig. A, and pl. 10, fig. 15). In the box cañon two to three miles below, the width of the river appeared to be much less, and, because of the precipitous walls, high water would make little change in width.
The two species of Ammospermophilus were seen at points only about 850 feet apart in a direct line. All the mammals of the Encelia association, as segregated here on the two sides of the river, were trapped at this station within one thousand feet of one another. The same situation obviously held at several other points along the Colorado River. The sharp separation of the ranges of nearly related vertebrates by a barrier of such narrow width is, to the best of the writer's knowledge, not known elsewhere in North America.
As to the opportunities for crossing the river by such individuals as might get to the water's edge, only speculation is now possible. At times of rising water, riparian mammals are undoubtedly often marooned upon islands and finally forced to swim or to take refuge on floating drift. Practically all the riparian species are known to be able to swim readily, and are probably in ways just mentioned frequently carried from side to side of the river.
The ability of typical desert animals to care for themselves if cast into the water is problematical, though a little experimentation would go far to proving the point one way or the other. Their powers in this line may be inferred to be limited because of the facilities normally lacking for putting such powers into practice. For instance, there is fair probability that a Reithrodontomys could safely cross a turbulent stretch of current, where a Perognathus would perish before any chance of reaching the shore.
Protracted observation along the river brings conviction to the observer that no animal of weak swimming powers is likely to survive many minutes of exposure to the main current. It is a fascinating diversion to watch the course of a stick or log adrift in the stream. Such an object pursues an exceedingly devious course. [[p. 104]] It may be carried close under the steep outside bank of an ox-bow swing, only to be directly thrown back towards the opposite shore. Stretches of rough water may be encountered where the object is swamped at the crest of every wave. Or, along rocky parts of the channel, swirls, large and small, arrest its passage. In the most violent of these eddies a twenty-foot log was seen to up-end and sink from sight, to reappear after its total submergence, a hundred yards down stream.
At times of falling water a great deal of drift lodges on mudbars and projecting reefs of rock. It is imaginable that drift logs might be reached by individuals, which freed again with subsequently rising water, would carry their passengers until lodged under favoring circumstances on the opposite side of the river. In the account of Neotoma intermedia desertorum, the only rodent of the Encelia association not checked by the river, it is suggested that in some such way passage was secured from the California to the Arizona side of the river. This wood rat now bids fair to occupy much appropriate territory in southwestern Arizona not previously possessing an associational homologue, that is a Neotoman representative. Neotoma may be looked upon as a more hardy and ecologically less specialized rodent than any of its associational companions. It is certainly much the largest, and is notoriously of aggressive disposition as a forager.
Of the eight species of delimited rodents, not one individual of the hundreds trapped was found on the "wrong" side of the river. As far as they went, then, our efforts furnished no evidence that even an occasional individual does get across. As already shown, there seems to be nothing to attract the upland rodents to the water's edge, so that possibility of securing safe transportation on a log or mass of drift is doubly remote. Now, supposing that a single individual did manage to reach the opposite shore, its species would not necessarily be established there. In most cases (not, however, with Peromyscus crinitus stephensi and Perognathus formosus) there is already established an associational homologue, with which even a whole family of the invaders would have to compete, with the chances at least as much against success as favoring it. Hybridization might occur, granted that no sexual antipathy arise, but, whatever the immediate results, it is the impression of the writer that swamping would eventually be likely to wipe out all trace of the invading species. This impression is admittedly based upon fragmentary data which has not been subjected to critical analysis. Whether or not Mendelian behavior in [[p. 105]] inheritance of characters obtains among the rodents here concerned is yet to be proven.
Suffice it to say that all the evidence at hand shows the Colorado River to have effectually blocked distribution, in the two directions concerned in the eight cases as listed. While this hindrance to distribution involves the species, it does so through its mechanical action upon the frontier individuals of each species. Hypothetically the invaders are severally hurled back or else destroyed outright.
The divergent characters displayed by the upland rodents of the two sides of the Colorado River are, in the mind of the writer, to be best explained on historical grounds. It is not necessary to believe that the specific characters concerned arose in the immediate vicinity of the river, though the circumstance of segregation alone is deemed by some to suffice as a cause of differentiation. The climatic features (zonal and faunal, as well as associational) are identical on the two sides of the river. Rather is it reasonable to presuppose separate and rather remote centers of differentiation, and convergent dispersal through time and space which brought the resulting types to the verge of the river, beyond which they were unable to spread.
It is possible that an arm of the sea continuous with the Gulf of California once extended northward into southern Nevada. A submergence of only 1,000 feet would divide the present desert areas of western Arizona and southern California into two peninsular land masses, which might have served as well-isolated centers of differentiation for various forms which later spread with the elevation of the land until their ranges abutted. Unfortunately for this suggestion, as I am informed by Professor John C. Merriam, geological evidence fails so far to show the existence of such conditions within Pleistocene or even Pliocene times. The suggested explanation must therefore be discarded in our dealing with the differentiation of present-day species and subspecies, especially since even the genera represented and as now restricted are not known to have evolved so early as Miocene.
But another process, recognizable far and wide in dynamic zoogeography, may be called into account without assuming any departure in the past from topographic and climatic conditions as they are today. Comparison of the fauna of the Lower Sonoran plains of south central Arizona with that of the Mohave desert plateau in the same zone, shows two prevalent character combinations among the nearly related component species. The mid-Arizona representatives are usually dark colored and large sized, the reverse appearing to hold in the majority [[p. 106]] of the Mohave desert forms. There is considerable floral difference in the two regions, and minor climatic differences are well known. Different environments thus impinge upon the animals in these widely separated centers, and more or less regular blending of conditions occurs between. Although every factor of environment may be identical immediately on the two sides of the Colorado River, the animals now there have undoubtedly descended from ancestral lines which have invaded the territory from the two opposite directions, bringing with them by inheritance the characters developed under the two different sets of conditions.
To express the idea otherwise, from each differentiation area there is an outwardly radiating dispersal of descent-lines, involving time as well as space. This dispersive process is going on now as it has through past time. The eastward-flowing tide of Mohave forms would only be arrested by an insuperable barrier, such as the Colorado River. The westward invading descent-lines from the Arizona center would proceed until stopped by the same barrier. Both sets of forms would find themselves along the Colorado Valley under the same associational, faunal and zonal conditions; but each set is continually receiving by the process of inheritance plus invasion the peculiar characters generated on its own side.
While the Colorado River probably lies in an intermediate position between the Mohave and Arizona faunas, the area of intermediate conditions of environment is probably relatively narrow. This very element of narrowness may be called in to account for the lack of modification displayed by the delimited species of the Colorado River frontiers, for example, in the case of Ammospermophilus.
Supposing now that the Colorado River does not serve as an insuperable barrier, nor ever has done so; invasion would have extended from one side to the other as far as associational, faunal or zonal barriers permitted. In animals of wide distribution, intergradation geographically between the remote extremes would in the end be expected to occur. The extremes would not then have differentiated so far, at least in quantity of each character developed, because of inheritance from the opposite type, again involving time and space, concomitantly. Subspecies would have resulted, instead of full species. This condition doubtless obtains in some of the birds, as well as in some of the rodents listed as being the same on the two sides of the river. Take, as an example, Dipodomys merriami merriami of south central Arizona, and Dipodomys merriami simiolus of the Mohave [[p. 107]] desert. Our Colorado River series is fairly intermediate between the extreme types, though the extremes are not so different as are Ammospermophilus harrisi harrisi and Ammospermophilus leucurus leucurus. As elsewhere explained, the river is believed to be not so much of a barrier to Dipodomys merriami as to the forms of Ammospermophilus.
The degree of hindrance, ranging from the condition, as in the case of Castor and Ondatra, where the river offers no bar to perfect freedom of crossing, to that where the river is an absolute barrier, as in the case of species of Perognathus and Ammospermophilus, accords so closely with degree of difference in characters developed on the two sides of the river, that adequate ground is afforded for the belief that intervention of barriers is a prime factor in the differentiation of species. And furthermore, it would appear that no two species, in birds and mammals, arise except through geographic segregation.
THE PROBLEM OF BARRIERS WITH REGARD TO BIRDS AND MAMMALS
The geographical range of any species of animal may be likened to a reservoir of water in a cañon. The confining walls are of varying nature. A concrete dam, absolutely impervious, may retain the water at one end. Along either side the basin's walls differ in consistency from place to place. The substratum varies in porosity, at some points impervious like the dam, at others permitting seepage of water to a greater or less distance from the main volume. The water continually presses against its basin walls, as if seeking to enlarge its area. And it may succeed in escaping, by seepage through such portions of its barrier as are pervious or soluble, or by free flow through a gap in the walls, if such offers. The area occupied by the water will extend itself most rapidly along the lines of least resistance.
Every species has a center or centers of abundance in which favoring conditions usually give rise to a rate of reproduction more than sufficient to keep the critical area stocked. A tendency to occupy a larger space results because of competition within the species, and individuals and descent-lines multiply and travel radially, extending those segments of the frontier where least resistance is offered. Such radial dispersal takes place slowly in some directions, more rapidly in others, according to the degree of passability of the opposing [[p. 108]] barriers. These barriers consist of any sort of conditions less favorable to the existence of the species than those in the center of abundance.
Theoretically, sooner or later and in all directions, every species is absolutely stopped. But as a matter of undoubted fact most barriers are continually shifting, and the adaptability of the animals themselves may be also undergoing continual modification; so that perfect adjustment is beyond the limits of possibility so long as topography and climate keep changing. The ranges of species may thus be constantly shifting. Descent-lines may move about repeatedly over the same general region, like sparks in the soot on the back of a brick fireplace.
Yet, in all of our studies, of but a few years' duration, the time element is reduced almost to a negligible quantity, and we may look upon the areas occupied by each species as, for the time of our observation, fixed. We are thus enabled to compare one with another, and because of the large number of the species, we can infer a good deal as to the nature of barriers in general as regards birds and mammals. It is even conceivable that with sufficient refinement in methods the inquirer may in time find himself able, from a comparative study of the ranges of rodents, for example, to establish the identity of all of the external factors which have to do with the persistence of each species; in other words to analyze the "environmental complex" into its uttermost elements as regards the existing species of rodents in their recent development.
The most obvious kind of barrier to distribution is that consisting of any sort of physical, or mechanical, obstruction. Such obstruction affects directly the individuals of a species encountering it, either by stopping their advance, or by destroying outright such as attempt to cross it. As barriers of this nature are to be cited land to purely aquatic mammals, and bodies of water to purely terrestrial, especially xerophilous, mammals. In each case width of the barrier has to do with degree of impassability. Oceans and continents are most perfect and affect a large proportion of the species. The comparatively narrow Colorado River is a barrier of the first rank, but only to a certain few desert rodents, as pointed out in a preceding chapter. Mechanical barriers, where they exist at all, are clearly recognizable.
It is to be observed, however, upon considering the birds and mammals of a whole continent, that by far the greater numbers of species are delimited in range irrespective of any direct dependence upon actual land and water boundaries; more explicitly, their ranges fall [[p. 109]] far short of coast lines. The barriers here concerned are intangible but nevertheless powerful. By their action the spread of species, genera, and families is held in check as surely as by any tangible obstruction.
By these invisible barriers the individual may not necessarily be stopped at all, as with animals of free locomotion; but the species is affected. For example, the mockingbird (Mimus polyglottos leucopterus) in its Californian distribution is closely confined to those parts of the state possessing certain definite climatic features; but vagrant individuals, especially in autumn, occur beyond the limits of these restrictive conditions. Carnivorous mammals are well known to be subject to sporadic wanderings on the part of individuals, but the species is kept in set bounds by some potent but invisible set of factors. The very fact that individuals are quite capable of temporarily transgressing these bounds and yet do not overstep them en masse emphasizes all the more the remarkable potency of this category of barriers as regards species and higher groups.
Our geographic studies lead us to designate among these relatively intangible barriers: (1) increase or decrease in prevailing temperature beyond certain critical limits, according to the species concerned; (2) increase or decrease in prevailing atmospheric humidity beyond certain limits; (3) modification in food-supply and appropriate breeding and foraging ground as regards the inherent structural equipment of each animal considered. In these three sorts of barriers will be recognized the zonal, faunal and associational delimitation as discussed in previous chapters.
CLASSIFICATION OF BARRIERS TO SPECIES
AS REGARDS BIRDS AND MAMMALSBarriers:
A. Tangible (mechanical)
(a) Land to aquatic species
(b) Bodies or streams of water to terrestrial species
B. Intangible (non-mechanical)
(a) Zonal (by temperature)
(b) Faunal (by atmospheric humidity)
(c) Associational
(1) By food-supply
(2) By breeding places
(3) By temporary refuges
(Each of these three with regard to the inherent structural characters of
each species concerned).
[[p. 110]] The above categories are believed to include all the factors commonly involved in the checking of the spread of species of birds and mammals. It is possible that inter-specific competition may sometimes occur where associational homologues meet. But even here it becomes a matter of relative associational fitness which determines supremacy and consequent ultimate limits of invasion of the forms concerned.
A mountain range is no barrier at all, per se, as frequently alleged. Only as it involves zonal or faunal barriers does it affect distribution. The same is true of a valley or a desert.
As far as contemplation of cases has gone, the writer's experience has led him to believe that the outlines of the ranges of all birds and mammals may be accounted for by one or more of the factors indicated in the above analysis. And as detailed knowledge of the facts of geographical distribution accumulates, the delimiting factors become more and more readily detectable. By such a study, of comparative distribution, it seems possible that the ranges of birds and mammals may become subject to satisfactory explanation. The instances included in the list of species discussed in the present paper, when considered in connection with many similar ones, point without exception towards the existence of the set of factors above specified as delimitors.
When considered in its historical bearing, the problem of barriers concerns itself intimately with the origin of species. It is believed by the writer that only through the agency of barriers is the multiplication of species, in birds and mammals, brought about.
* * * * *