
http://people.wku.edu/charles.smith/biogeog/SIMP1950.htm
History of the Fauna of Latin America
by George Gaylord Simpson (1950)
Editor Charles H. Smith's Note: Original pagination
indicated within double brackets. My thanks to American Scientist
for permitting the reprint. Citation: American Scientist 38 (1950):
361-389.
[[p. 361]] It is
odd that there should be a Latin American fauna, a broad unit
that can be roughly designated by such a term as "Latin American," defined
by human linguistics and culture. The animals inhabiting this area can
hardly have foreseen that the dominant languages of the twentieth century
would here be Spanish and Portuguese or that the European cultural elements
imported here would come mainly from Latin Europe--from Spain and Portugal
and also, in considerable measure, from France and Italy. Nevertheless,
there is a characteristic fauna that coincides approximately with Latin
America and that differs in some major traits from the fauna of English-speaking
America. The coincidence is not precise. The fauna of northern Mexico,
although transitional in some respects, is more nearly allied to that
of the United States than to that of most of Latin America. Exact correspondence
of native fauna and imported culture would be a miracle, and it is still
a wonder that the equivalence is as close as it really is. There is, indeed,
a common factor that removes the correspondence from the realm of pure
coincidence. The demarcation both of faunas and of cultures has been influenced
by climatic factors.
Radical differences in the two major faunal realms of the Americas were noticed by the early explorers and are still obvious enough to the modern traveler. A New Englander traveling in Brazil does not need to be a zoologist to observe that the animals of that country are at least as exotic as are the speech and customs of its human inhabitants. He will see some animals which, although subtly different, look reasonably familiar to him: deer, foxes, field mice, squirrels, rabbits, and a few others. More will be completely new to him outside of zoos: peccaries, tapirs, jaguars, kinkajous, guinea pigs, agoutis, capybaras, armadillos, tree sloths, monkeys, and a host of others.
The distinction is not confined to mammals, a few of which have been named, but extends to all sorts of animals. The river rays, the lungfish, the piranhas, and many other freshwater fish are strange to northern eyes. The frogs and toads may look familiar at first sight, but on closer [[p. 362]] inspection such forms as the Surinam toad, incubating the young in its back, and, indeed, almost all others will be found distinctive. Among turtles, the matamata and other side-necked forms are obviously exotic, and so are the boa constrictors and numerous other snakes. The ostrich-like rheas and the wing-clawed hoatzins are only two among the numerous purely Latin American birds. Additions to the long roster could be provided among earthworms, insects, and innumerable others.
The fauna is by no means uniform all over Latin America, but it is evident, first, that broadly similar faunal characteristics do appear throughout most of this great area and, second, that these characteristics distinguish the region sharply from non-Latin North America or any other continent. These facts led the students of animal distribution in the latter half of the nineteenth century (the Sclaters, Wallace, Beddard, and Lydekker, to name some of the more eminent) to set Latin America aside as the Neotropical Region or Neogaea. In the classical arrangement, this region includes all of South America (even those parts decidedly nontropical), Central America, tropical Mexico (but not the temperate central plateau and northern Mexico), and the West Indies. Although the boundaries are sharp and definite on the map, it was of course recognized that the line in Mexico is not awesomely respected by the animals and that it really lies within a broad transition zone. Some mainly neotropical animals, such as the peccaries or jaguars, range into the United States, far north of the map line. Some mainly nearctic (non-Latin North American) mammals, such as shrews, similarly range well south of the line. It has also long been recognized that the West Indian fauna is not typically or fully Latin American or neotropical. It is impoverished relative to the mainland, has its own peculiar forms, and has a few special resemblances to the North American fauna.
In spite of some ambiguities, the Neotropical Region does have a well-established clarity and validity in the zoogeography of the world as it exists today. This static picture is, however, the result of a long and dynamic historical process. The nature of the fauna in what is now designated as the Neotropical Region has changed radically during geological history. Different parts of that region have not had the same history. Faunal resemblances and distinctions have sometimes been greater, sometimes less than they are today. Boundaries of faunal assemblages have not remained in the same place. Recent neotropical elements may be old in that region or may be latecomers with their historical faunal associations elsewhere.
Until relatively recently, geologically speaking (well into the Pliocene), Central America had faunal affinities almost exclusively with North America and hardly at all with South America. There was then no Neotropical Region in the present sense. American camels, tapirs, and other animals now exclusively neotropical were pan-American in the Pleistocene and exclusively (in this hemisphere) North American before that. They are surely not neotropical in the historical sense, or in the same sense as, say, armadillos, which arose in South America [[p. 363]] and were until the late Tertiary confined to part of the Neotropical Region as now delimited.
In a historical view, then, the Neotropical Region ceases to be clear and consistent. Its fauna is not a coherent unit from this point of view, and its boundaries are not even approximately constant. In the historical study the static concept must be abandoned. The region and its fauna cannot be taken as either definite or invariable, but must be analyzed through a long series of shifts and fluxes.
Faunal Strata
In any given region, the various groups of animals will be found to have occupied that territory for different lengths of time. In general, the longer the group has been there, the more peculiar it will be to the local scene and the more strongly differentiated from relatives living elsewhere. The results of this process are particularly striking in South America. Armadillos and sloths belong to an order (Edentata) known nowhere else (aside from marginal spread into North America). Differentiation has proceeded so far in that region that there is now no clear trace of special relationship to animals of other regions, aside from the broadest fact that these edentates are placental mammals. The monkeys of South America are quite distinct, as a group, from those of the Old World and yet clearly related, belonging to the same order, Primates. They show an intermediate degree of regional differentiation. The field mice of South America, although distinctive as to species and, usually, genera, are closely related to those of North America, belonging not only to the same order (Rodentia) but also to the same family (Cricetidae) and even to the same subfamily and tribe. They have a low degree of regional differentiation.
The fossil record clearly confirms the fact that these varying degrees of differentiation are correlated with the time when the ancestors of these animals were emplaced in South America: edentates, high differentiation, in the earliest Cenozoic; primates, medium differentiation, in the mid-Cenozoic; cricetids, low differentiation, in the late Cenozoic. The three groups represent three readily distinguished faunal strata. The faunal history of South America has clearly been episodic.
The faunas of all regions are apparently stratified and their histories episodic in much this same way. On most continents, however, episodes of emplacement of new groups have been so frequent and so scattered in time, and their isolation from allied groups of other regions has been so imperfect, that the stratification is highly complex and the distinction of various strata is blurred. Only in South America and in the present island continent, Australia, are the strata relatively few in number and, as a rule, sharply and clearly separable. This clarity is a result of the physical history of the continent. Stratigraphic and paleontological evidence agree in clearly demonstrating that South America was isolated by sea barriers from all other continents from about the beginning of the Tertiary until near its end (probably early Paleocene to late Pliocene). During this long span, on the order of 70 million years, its [[p. 364]] animals were genetically isolated from all other continental faunas, and introduction of new stocks was, although not completely prevented, extremely restricted.
Some complications exist in each case, but in broad lines the South American mammalian fauna can be analyzed into three strata. The oldest stratum, which may be called that of the "ancient immigrants,"

includes groups that reached South America before its complete isolation from the rest of the world, in latest Cretaceous or earliest Tertiary times, and the numerous and varied descendants of these ancient stocks. The second stratum, that of the "old island-hoppers," includes only two groups, old native rodents and monkeys, which reached South America while it was isolated. The third major stratum, of "late island-hoppers" and "late immigrants," with considerable complexity of detail, includes a [[p. 365]] great variety of animals that reached the continent in the late Tertiary and the Quaternary, shortly before and during its re-connection with North America. The three mammalian strata are shown in more detail in Table I, and their correlation with geographic events is diagrammatically suggested in Figure 1.
For animals other than mammals, the fossil evidence is relatively poor. Most of the vertebrates have more or less clear indications of the same threefold stratification, with some probable blurring of the picture for groups less rigidly restricted by a sea barrier (such as some birds and probably some fishes). Among freshwater and terrestrial invertebrates, the fossil record of which is almost nil, there may be an added complication in the form of faunal strata still older than the oldest for mammals and most other vertebrates. On the whole, however, the
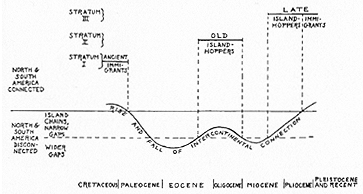
Fig. 1. Relationships between faunal strata in South America and the rise and fall of intercontinental land connections. The graph is diagrammatic only, as the history of the connection was not so simple as shown, and the sea barriers shifted in position.
three designated strata are pervasive in historical interpretation of the entire fauna.
These faunal strata can be designated as such and thus dated only with respect to the geographic unit of South America and by virtue of its Tertiary isolation. In Central America, for instance, much the same groups of animals occur, but their time relationships and their associations in strata are quite different with respect to the region occupied. The oldest mammalian faunal strata in Central America include what are in South America the old island-hoppers and various of the late island-hoppers and immigrants. The old immigrants of South America, forming the oldest stratum there, belong to what is generally speaking the youngest stratum in Central America. There are profound differences in regional history in parts of what now is the broadly unified single Neotropical Region.
[[p. 366]] The Problem of Basic Mammalian Faunal Types
A fundamental problem regarding the Latin American mammalian fauna arises at the very beginning of its history. When the Age of Mammals began, with extinction of many groups of Mesozoic reptiles and the beginning of the rise of mammals to dominance in land faunas, the mammals were already, in fact, an old group. Although still small, obscure, and relatively unvaried, the major lines of mammals had then already split into two basically distinct, primitive groups: marsupials and placentals or, technically, Metatheria and Eutheria.
Africa, Europe, Asia, and North America then formed an essentially continuous land mass, a World Continent. The continuity has been interrupted at various points from time to time and there have been innumerable faunal changes and marked regional differentiation in various groups, but the mammalian faunal type was then and remains today fundamentally the same throughout all parts of the World Continent. Marsupials were present at the start (at least in North America and Europe, and probably throughout), but they were of minor importance. The major ecological roles, and specifically those of carnivorous and herbivorous types, were played, as they still are, by placentals.
In Australia, probably then already an island continent (Notogaea), and in South America, then or shortly thereafter becoming an island continent (Neogaea), the basic faunal types with which the Age of Mammals began were radically different from the World Continent type. In Australia all the mammalian ecological roles were played by marsupials. (Such early faunas have not, in fact, been found in Australia, but this is a probable and generally accepted inference from the evidence of later faunas.) In South America, the herbivores were mainly placentals, but the insectivorous, gnawing, and carnivorous types were marsupials.
Early attempts to explain this extraordinary anomaly between the South American island continent mammalian faunal type and the World Continent type usually involved the theory that South American marsupial carnivores were derived from Australia. (Why the carnivores of Australia were marsupials rather than placentals is another fundamental problem not here under consideration.) Among several serious objections to this idea is the extreme improbability that marsupial carnivores would spread from Australia to South America unaccompanied by their ecological associates and prey, the Australian marsupial herbivores. Other possible sources of mammalian faunas all had placental carnivores and placental herbivores, and equal difficulty is encountered in explaining the spread to South America of these herbivores without their associates, the placental carnivores. That the carnivorous and herbivorous elements in the first, balanced, basic South American mammalian fauna should have come each separately and from a totally distinct source is simply incredible.
An alternative view is that the basic South American fauna did have an essentially unified geographic origin, all coming from (or at any rate being connected with) the World Continent fauna. In that case, [[p. 367]] however, it would appear that placental carnivores must have been included but must almost immediately have become extinct in South America. Such extinction would seem inherently improbable, because in the established cases of competition between marsupial and placental carnivores, the placentals have survived and the marsupials have become extinct. The placental dingo survived in Australia and its marsupial analogue, the thylacine, became extinct (except on the separate island of Tasmania which was not reached by the dingo). The last South American marsupial carnivores disappeared as placental carnivores (of the latest major faunal stratum) arrived. It is an additional difficulty that specialized marsupial carnivores are quite unknown from any age on the World Continent and probably never occurred there. Similarly, placental carnivores are totally unknown in the early South American faunas, and extinction so rapid and complete as to eliminate all traces in the now rather abundant early Tertiary fossil collections seems improbable, although not impossible.
These objections largely disappear, and a reasonable solution of this long-disputed problem can be suggested, if one gives consideration to the relatively undifferentiated and primitive mammals of the earliest Tertiary, rather than thinking in terms of the more familiar, sharply distinct and specialized later forms. The basic World Continent fauna, at its very beginning, did not include radically differentiated placental carnivores and herbivores. These two later ecological types were then ancestrally represented by one basic stock, to which I have elsewhere applied the name "ferungulate." This stock was rapidly becoming considerably varied and apparently included some rather more herbivorous and some rather more carnivorous forms, but as a whole it was omnivorous and all its members were still closely similar. Among the fossils of this group, some are classified as Carnivora and some as Condylarthra (primitive ungulates), because of our knowledge of the subsequent sharp ordinal distinctions of their descendants. If we did not know their later history, the early forms would certainly be placed in the same order and very likely in the same family or even smaller group. For instance, Protogonodon, an early Paleocene form classified as a carnivore, and Desmatoclaenus, also early Paleocene, classified as an ungulate, are far more alike than numerous pairs of recent genera referred to one family or subfamily, say Mustela, the weasel, and Gulo, the wolverine, both commonly placed in the subfamily Mustelinae.
It was this more generalized ferungulate stock, and not any later and then truly distinctive placental carnivores and herbivores, that figured in the beginnings of the oldest faunal stratum of South America. On the World Continent, this stock was associated with likewise primitive marsupials, which also lacked as yet any distinctive specializations as carnivores but were equally capable (as the event proved) of such a development. In the World Continent, after it was cut off from South America, specialized carnivores arose among the ferungulate lines, and their occupation of this ecological specialty impeded the rise there of marsupial carnivores. In South America, likewise after separation [[p. 368]] from the World Continent, marsupials more rapidly developed carnivorous types. This in itself would tend sufficiently to inhibit the rise there of placental carnivores among the ferungulates, which rapidly developed a great variety of types all more or less herbivorous.
Thus there was no competition between marsupial carnivores and placental carnivores, as such, and probably only marginal competition between marsupials and placentals as a whole, but only a parceling out of the various ecological zones, which happened to receive different occupants in the World Continent and in South America. Why the ferungulates evolved more rapidly into specialized carnivores on the World Continent and the marsupials (didelphoids) in South America is not evident. It is, nevertheless, plausible that this could occur, and the hypothesis that it did occur seems to be the only one that reasonably accounts for these differences in basic faunal type.
There may be some remaining objection that placentals in general are superior to marsupials and would tend, in South America or elsewhere, to occupy any and all ecological niches, including those of carnivores, at the expense of the marsupials. This possible objection really has little or no force. Present evidence is that the placentals are not more advanced derivatives from backward or older marsupials, as was commonly believed in the nineteenth century and is still occasionally stated in textbooks. Placentals and marsupials seem rather to represent a basic dichotomy of the main mammalian stock and to have been about equally progressive and adaptively efficient when they arose. Primitive forms of the two major branches apparently lived together in ecological equilibrium, and even today the opossums of North and South America remain abundant and are eminently successful in holding their own in the midst of the dominant placental faunas. Ecological incompatibility between marsupials and placentals seems to arise only when late and narrowly specialized forms come in contact within essentially the same ecological niche, a situation in which any two groups, and not only marsupials and placentals, become ecologically incompatible. Even on this score, the extinction of South American and Australian marsupial carnivores in competition with placental carnivores gives evidence not particularly that placentals are superior to marsupials but that the late World Continent groups had become competitively superior to those of the island continents. In South America, the old native placental groups were also decimated when they came into contact with placentals from the World Continent. They fared no better than the marsupials of similar geographic history.
Origins of the Oldest Faunal Elements
Probably the most disputed single question of Latin American zoogeography has been the geographic origin or, at least, connections of the older faunal elements. The principal question raised has concerned possible direct relationships among the southern continents, between South America and Australia, on one side, or Africa, on the other. The various postulates include transoceanic land bridges or continents, [[p. 369]] land connections by way of Antarctica, or early continental union followed by fracturing and drift (Wegenerian) to the present widely separated positions of the three continents. A summary of the truly voluminous and polemic literature of this subject is outside the scope of
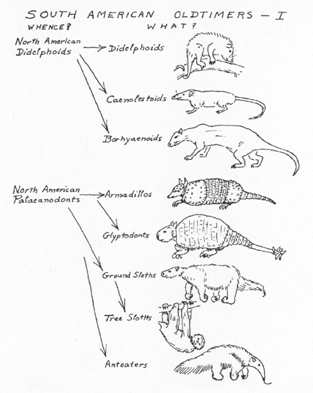
Fig. 2. Marsupials and edentates, evolved in the oldest mammalian faunal stratum of South America, and their possible origins. (Not drawn to scale.)
the present study, but the nature of the older mammalian faunas and the evidence these give as to their origin will be briefly reviewed.
The major elements among the mammalian old immigrants and their possible origins are as follows (see also Figs. 2 and 3).
[[p. 370]] MARSUPIALS. These were already quite varied in the older Tertiary, with primitive, more or less insectivorous types, rodent-like types, and marsupial carnivores. A possible common origin of all would be in a varied assemblage of relatively unspecialized marsupials, didelphoid or
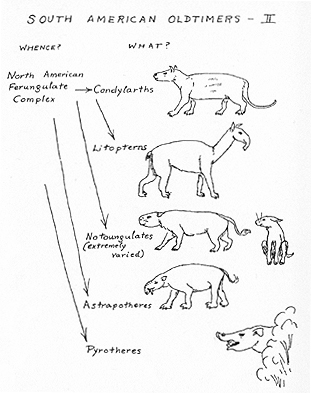
Fig. 3. Ungulates of the oldest mammalian faunal stratum of South America, and their possible origins. (Not drawn to scale.)
extremely primitive dasyuroid in general character. Such an assemblage is known from the late Cretaceous of North America, where primitive marsupials (opossums) also occurred through the Paleocene and later. Similar forms occur doubtfully in the Paleocene and surely in the Eocene of Europe. Elsewhere the evidence is purely negative.
[[p. 371]] EDENTATES. The oldest South American faunas include only armadillos, of primitive type, among edentates. The other groups appear, evidently by evolution within the continent, during the early and middle Tertiary: glyptodonts and ground sloths by late Eocene, anteaters in the Miocene, tree sloths (with practically no established fossil record) by rather minor differentiation from the less specialized ground sloths sometime around the mid-Tertiary. The only firmly established special relationships with any non-South American mammals are with the Palaeanodonta, a group more primitive in general character than any of the South American edentates and known only from North America (late Paleocene to middle Eocene, with a probable more specialized offshoot in the early Oligocene).
CONDYLARTHS. These most primitive of ungulate herbivores were fairly common in the Paleocene-Eocene faunas of South America and some lingered on into the Oligocene (or possibly the Miocene). The same order was important in the basic World Continent fauna. It occurs abundantly throughout the Paleocene and Eocene of North America and some North and South American genera are suggestive of special relationship.
LITOPTERNS. This order, known only from South America, was common and varied in early faunas and persisted, in decreasing variety, into the Pleistocene. Early forms are near the condylarths, to such an extent that the litopterns might be considered merely as surviving and diversely specialized condylarths. They seem to have originated in South America from the South American condylarths, and therefore to have the same source as the latter.
NOTOUNGULATES. This great order of hoofed herbivores constitutes the bulk of all the earlier South American faunas and continues, with radically decreasing variety, into the Pleistocene. It suggests a group similar in origin and parallel in history to the litopterns. An odd anomaly arises from the discovery of a notoungulate in the late Paleocene of Asia and a related form in the early Eocene of North America, the only occurrences of the order outside of South America. Origin in Asia and migration to South America by way of North America are suggested by the face of these facts, and this remains a possibility but is rendered rather improbable by various considerations. Early Eocene is too late a time for entry into South America. The most primitive South American forms are slightly but definitely less specialized than those known from Asia and North America. Another possibility is that the latter were strays from South America. The question cannot now be resolved, but a faunal connection of one sort or another is indicated with the northern continents.
ASTRAPOTHERES. Although never highly varied, these aberrant hoofed herbivores appeared early in South America and persisted into the Miocene. They probably arose in that continent and have no evident close or special relationships with other known groups, except that they probably originated ultimately from condylarths or most primitive ferungulates in a broader sense. They could have some collateral [[p. 372]] relationship or, at least, functional parallelism with archaic World Continent forms like the pantodonts and uintatheres, known in particular abundance from North America but also spreading to Eurasia.
PYROTHERES. These odd, superficially mastodon-like ungulates were never much varied or particularly common, as far as known, but they appear in early Eocene faunas and persist into the Oligocene. They are known only from South America and probably arose in that continent. Special relationships are not known, but in broad terms differentiation from the basic ferungulate complex is indicated, more or less as for the astrapotheres. True relationship to the Proboscidea has been suggested but now seems untenable except as the Proboscidea may also represent a separate branch from a part (paenungulate) of the ferungulate complex.
The data on the very earliest Tertiary mammals of South America and of the world as a whole are so scanty that tracing of exact lineages cannot be expected and is not, in fact, possible at present. In broad terms, this assemblage certainly suggests derivation from the World Continent fauna and could not be derived from Australia unless it is assumed, quite gratuitously and contrary to such evidence (all indirect) as exists, that Australia did once have a basic fauna of World Continent type and later eliminated almost the whole of this fauna. Within the World Continent, there is absolutely no evidence of any special relationship of Africa with South America. This is partly a negative conclusion, because Paleocene mammals are unknown from Africa, but it would, again, be wholly gratuitous to assume that relationships existed and left no trace in the late Eocene and subsequent African faunas.
Such definite evidence of relationships as exists all points most nearly to North America. Every South American stock is related at least as closely to one known from North America as to any other known group outside of South America, and the edentates have no established relationships except with an exclusively North American group. In view of these relationships and of the fact that South America is now geographically closest to North America (and nothing impels the belief that this was untrue at the end of the Cretaceous), the tendency of so many students to look elsewhere for the main geographic relationships of the old South American fauna almost smacks of a preference for the unlikely over the obvious.
The question remains whether the World Continent fauna might not have been derived from South America, as the great Argentine student, Ameghino, insisted, rather than the other way around. In the exact sense of Ameghino, who believed that later specialized orders and families originated in South America and spread hence over the rest of the world, this certainly is not true. In the sense that some elements of the most basic World Continent fauna might have become differentiated in South America while that continent was united to North America and hence in a sense part of the World Continent, the possibility remains but it seems quite improbable. The old South American faunas seem to represent diversification on that continent from a sampling of the World Continent fauna that was partial only (no insectivores, no early primates, no early [[p. 373]] rodents, no differentiated creodonts, only highly aberrant paenungulates). No forms conceivably near the ancestry of any World Continent lineages have ever actually been found in the old South American faunas.
Clearly inconclusive, the evidence does suggest North America as the probable source of the old South American fauna. It does not exclude, but definitely does not support, other possibilities. This conclusion finds further, indirect but strong substantiation from the evidence that South America was connected with North America toward the end of the Age of Reptiles. Although opposite statements may still be found in the literature, it is at present recognized by the most competent specialists on these faunas that the late Cretaceous reptiles of South America are, as a group, more like those of North America than like any known faunas elsewhere in the world.
The Old Island-Hoppers
The origin of the second faunal stratum of South America, although less discussed as a separate problem, has been in many ways even more puzzling. This stratum comprises the old native rodents, which are the so-called South American hystricomorphs, and the neotropical primates (see Fig. 4). These rodents first appear in the record in beds somewhat uncertainly correlated as early Oligocene, and primates are first known from later beds, considered with similar uncertainty as late Oligocene. Both occurrences are marginal on the continent (in Patagonia) and do not exclude the possibility that the groups had been for some time in more central or northern parts of the continent, where adequate faunas of appropriate age have not been discovered. It is, however, improbable that emplacement of these groups in South America was long prior to the Oligocene and it seems almost certain that their ancestors were not among the early immigrants. There is no evidence that entry of the two groups was absolutely simultaneous, and, indeed, it probably was not. Both, however, entered South America at about the same time, roughly midway between the two major immigrations; the two have become comparably differentiated there, and it is justified to consider them broadly as of the same faunal stratum.
Both these groups have repeatedly been cited as indicating faunal relationships with Africa. The neotropical monkeys, although distinctly definable as a group, resemble the Old World (including African) monkeys and are on a similar evolutionary level. The old South American rodents also show resemblances to some Old World rodents (porcupines figure in both regions, for instance), and among them are some, in the general group of the hutias, degus, tucutucus, and spiny "rats" (not true rats), that particularly resemble some African rodents, the cane and rock "rats" (also not true rats).
Recent studies seem rather conclusively to controvert these apparent African affinities. The South American primates, including the earliest forms, are in some respects more primitive than the Old World forms of similar or later age and in some respects differently specialized. The [[p. 374]] former characters seem to prohibit derivation of the New from the Old World forms, and the latter seem to exclude filiation in the other direction. There is a strong suggestion that the New and Old World monkeys
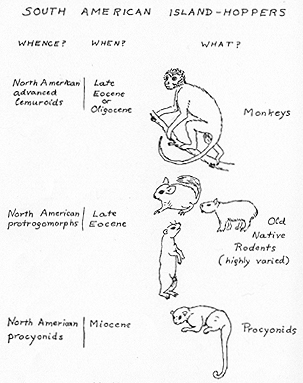
Fig. 4. Derivatives of island-hopping immigrants into South America, and their probable origins. The monkeys and old native rodents form the second or intermediate mammalian faunal stratum. The procyonids (allies of the raccoon) may be considered rather as forerunners of the last, relatively complex faunal stratum. (Not drawn to scale.)
represent geographically separated parallel developments from a more remote and primitive (technically prosimian) ancestry. About twenty-five years ago, the late J. W. Gidley suggested that neotropical monkeys [[p. 375]] might be derived from a group of prosimians (Notharctinae) relatively abundant in the Eocene of North America, and the Old World monkeys from Old World allies of this group. Too little attention has been given to this suggestion, but recent study is adding some evidence in its favor and it is now the best working hypothesis as regards the New World forms, at least.
It has been suspected from time to time that the history of the New and Old World "hystricomorph" rodents might be a similar case of independent, parallel development from allied New and Old World groups of primitive Eocene rodents, but concrete evidence has been scanty. Quite recently (in 1949), in describing relatively complete material of the oldest and most primitive known South American rodent, A. E. Wood has found positive evidence for their derivation not from a distinct hystricomorph stock but from a widespread World Continent group of most primitive rodents (Ischyromyidae, sensu lato). Among these, he finds particular resemblance to some North American forms (especially the mid-Eocene Reithroparamys).
There is, then, good reason to believe that the idea of special African relationships for these groups, old South American rodents and monkeys, is incorrect and that both are of North American origin. The evidence is, indeed, better than for the older immigrants, for in each case a possible North American ancestry can be rather closely designated among well-known groups.
It seems quite clear that these groups did not follow a land bridge from North America. When they entered South America, North America was swarming with rapidly progressive mammals of other types, notably many placental carnivores and a variety of perissodactyl and artiodactyl ungulates. It is incredible that an open migration route existed without any effect other than spread southward of just two stocks, possibly only a single introduction in each case. Entry was almost certainly by waif dispersal over what I have called a "sweepstakes route." Both groups, initially small and probably arboreal animals, are ecological types especially apt for such dispersal. Geological evidence in Central America and northwestern South America perfectly fits this picture. During late Eocene and Oligocene, the pertinent times in this connection, there was clearly a series of seaways between North and South America, interrupted by a series of islands. The seaways would bar any extensive faunal interchange. The islands would facilitate overseas spread of a few special groups, literally island-hoppers. Although it would, of course, be possible to postulate a similar island chain elsewhere, to Africa, for instance, no evidence known to me really suggests this and the postulate is unnecessary and unsupported.
The immediate source of the immigrants would, of course, be Middle America or what is now tropical North America, and not the region from New Mexico to Montana where early Tertiary faunas are now known. Transition from the archaic ancestral groups (prosimians for the monkeys and protrogomorphs [of Wood] for the rodents) to the characteristic Latin American groups (ceboids and "hystricomorphs" or, perhaps [[p. 376]] better, caviomorphs, respectively) doubtless occurred in the paleontological terra incognita of early Middle America. Some basic diversification of these groups may also have occurred in Middle America and have been under way even before they island-hopped to the southern continent.
Development of the Native Fauna
The oldest reasonably well-known South American mammalian faunas (Eocene of Patagonia) are dominated by the typical and highly varied native ungulate herbivores of the Order Notoungulata, which includes nearly half of the known genera of that time. The rest of the known fauna is divided about equally among condylarths, litopterns, edentates, and marsupials (groups characterized on previous pages). Pyrotheres and astrapotheres were present and are striking animals, but were quite minor elements in the total fauna.
Until the late Pliocene, the faunal composition changed rather steadily but with few really profound modifications. The condylarths dwindled and finally became extinct, with ecological replacement by their collateral descendants, the litopterns. The notoungulates, exuberantly varied in the Eocene, continued in force but show a steady decrease in variety as primitive and intermediate lines were weeded out and a smaller number of more sharply distinct and specialized lines continued. On the other hand, the edentates, relatively little varied in the Eocene, expanded steadily into the Miocene when, in variety of genera, they constituted about a third of the known faunas. Pyrotheres and astrapotheres died out, the former in the Oligocene and the latter by the end of the Miocene.
The most noteworthy change before late Pliocene followed the appearance of the second faunal stratum, that of the old island-hoppers. The old native rodents expanded steadily and greatly into the Pliocene, where they include about a third of the known mammalian genera. Similar, but less intense, expansion of the primates may be postulated, although they are absent in most of the known fossil deposits, which generally represent facies unsuitable for this predominantly tropical and arboreal group. The broader lines of this long faunal development are seen in Figure 5.
The greatest interest of the phase of South American faunal history while the continent was an island lies in the fact that it is a sort of large-scale natural experiment in evolution. The old immigrants rapidly occupied the large and varied continent, and the faunas then evolved there in dynamic equilibrium, undisturbed by wider genetic interchange or by irruptions from without other than those of the old island-hoppers--and this exception is itself so relatively simple and analyzable that it almost ideally exemplifies the process and consequences of single major additions to an evolving fauna.
The first striking evolutionary phenomena illustrated are those of "explosive" or "eruptive" evolution and adaptive radiation. The poorly known Paleocene faunas and, particularly, the better known early [[p. 377]] Eocene faunas are at a most active stage of this process. Everything here indicates the culmination of an exceptionally rapid and burgeoning expansion of the mammalian fauna into a great variety of ecological niches which were, before this episode, empty or nearly so. The number of separate groups, from generic to family levels, at least, is exceptionally large. Intergradation between lines later widely distinct is still common. There seems to be considerable overlapping and even duplication in ecological types within the same or adjacent local faunas. Variation within specific populations is often exceptionally great, a basis for rapid diversification speciational in pattern, and an indication that marked segregation of characters and specialization of adaptive type are under way but still incomplete.
Disappearance of intermediate types and fixation of a smaller number of well-defined groups each with a characteristic, separate, and
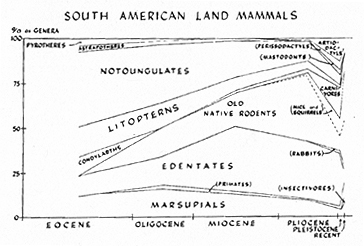
Fig. 5. Development of the land mammals of South America. For each epoch, the composition of the fauna is shown in terms of percentage of known genera belonging to the various orders of mammals.
progressively specialized adaptive and ecological status were processes evident through the later Eocene and Oligocene. This process was essentially completed in the Miocene as far as the descendants of most of the ancient immigrants are concerned, although the old native rodents and some of the edentates were then still in an expanding phase. The result of the process is a sort of parceling out of the available ways of life among the various stocks of mammals, an adaptive radiation.
In the meantime, on the World Continent even more varied ways of life were being occupied by adaptive radiation, or a more complex, interlocking series of over-all and local adaptive radiations, going on without contact with the South American fauna. When World Continent and South American lines became specialized for similar ecological roles, they also came to resemble each other functionally and morphologically. [[p. 378]] Parallel and convergent evolution was thus illustrated on a large scale. Another independent radiation in Australia produced another set of parallel and convergent types. Sometimes the separate lines departed from the same more or less remote common ancestry and evolved in close parallel, as in the case of the carnivorous marsupials of South America and of Australia. This process produces closest resemblance, a fact responsible for long debate on the affinities of these marsupial carnivores and for former insistence on the part of some students that South America and Australia must somehow have shared advanced and specialized marsupials and not merely the remote and primitive ancestors of these.
In other cases quite different groups have evolved toward similar adaptive types, a process producing less complete, convergent resemblance, as between the marsupial carnivores and the World Continent's placental carnivores. The two processes of parallelism and convergence intergrade and cannot be sharply defined in given cases. The striking similarity of some North American horses and some South American litopterns, evolving independently from a more or less remote condylarth ancestry, is a case in point. It can be interpreted either as straight parallelism from the common ancestral condylarth stage or as divergence, origin of the different orders Perissodactyla and Litopterna, followed by convergence between lineages in two families of these orders, Equidae in the first and Proterotheriidae in the second.
The faunas well illustrate the limitations of these processes, which produce similarities of various degrees but apparently never, even in cases of close parallelism, produce real identity, in part or in whole. In the flesh, a superficial or distant observer might well have confused Diadiaphorus, a South American litoptern, with Miohippus, a North American horse, but no competent anatomist would mistake any tooth or bone of one for the other.
Another interesting point is that such developments were not necessarily synchronous in the two cases. Thoatherium, a litoptern, was completely one-toed in the early Miocene. Horses did not become one-toed until the Pliocene, and even today Equus is less advanced than Thoatherium was in this respect (see Fig. 6). Some lines of notoungulates in South America had very high-crowned, complexly crested, cement-covered grazing teeth in the early Oligocene; horses did not reach a comparable stage until the late Miocene. These examples illustrate, by the way, that South American animals were not altogether less progressive or more slowly evolving than those of the World Continent, as has sometimes been supposed.
In many cases convergence was quite incomplete or would involve only a particular functional resemblance and not a close equivalence of total ecological status. Glyptodonts, for instance, the rigidly bulky cousins of the armadillo, seem to have been grazing forms and so have this functional resemblance with various ungulate grazers, but even their teeth are built on a plan wholly different from that of any ungulate. The glyptodonts as a whole can hardly be compared with any ungulate, [[p. 379]] or indeed with any World Continent animals. Such forms as some litopterns and some horses may be considered ecological vicars in their respective areas, but others, like the glyptodonts, are ecological uniques.
Late Faunal Mixture and Its Outcome
In the World Continent, there was a radical turnover in faunal type rather early in the Tertiary, mainly during the Eocene. In terms, for instance, of percentage composition of the mammalian faunas by orders or suborders, the difference between a Paleocene and an Oligocene fauna in North America is striking and almost absolute. Changes in this respect, of broad lines of faunal composition, have been relatively slight since the Oligocene. The modern faunal type was beginning to appear and to replace the oldest type even in the Eocene, thus here aptly called "Dawn of the Recent." Not so, however, in South America. There, as we have already seen in passing, change was gradual and involved no really fundamental upset of faunal type into the Pliocene. The mid-Pliocene fauna, in most of its broadest features, was not radically unlike a Paleocene fauna in spite of very pronounced advancing specialization in most of the orders and the Oligocene insertion into the fauna of two new orders (Primates, Rodentia).
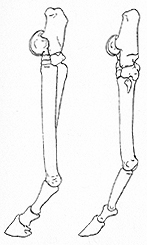
Fig. 6. Side views of hind feet of the modern horse, Equus (left), and of an extinct South American pseudo-horse, the litoptern Thoatherium (right), to show convergence in foot structure of these two one-toed mammals. Note vestiges of side-toes, larger in the true horse than in this pseudo-horse. (Not drawn to scale; Equus is larger than Thoatherium.) Redrawn after W. D. Matthew.
A change like that going on in the Eocene on the World Continent also occurred in South America, but at a greatly later date, in the late Pliocene and Pleistocene (Fig. 5). Its cause was the rise of the Central American bridge and the consequent irruption into South America of many derivatives from the fauna of the World Continent. This third broad faunal stratum did not come in all at once, in a single wave. Already in the late Miocene a few northern forms appeared, small arboreal placental carnivores more or less related to the raccoon. Not long thereafter, apparently in early Pliocene times, some South American animals, ground sloths, reached North America. These forerunners do not seem to indicate a continuous land connection but probably utilized [[p. 380]] the island chain, gaps in which were closing progressively as the Central American and northwestern South American regions rose relative to sea level. The exact moment when the bridge became complete is not
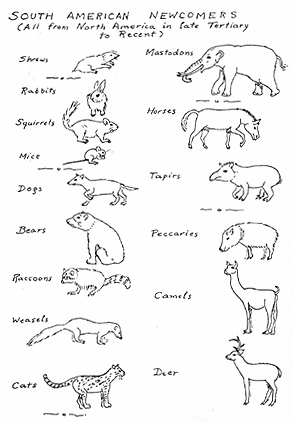
FIG. 7. Representatives of the principal families of previously North American mammals that invaded South America late in the Cenozoic, and which form the youngest mammalian faunal stratum there. (Not drawn to scale.)
[[p. 381]] established, but this probably occurred during the age called Chapadmalalan in South America and Blancan in North America, placed by some authorities as latest Pliocene and by others as earliest Pleistocene. Even then the exchange was at first rather limited in scope and the full surge of intermigration did not occur until somewhat later, in unequivocally Pleistocene times. Soon or late, at least fifteen (possibly sixteen) families invaded South America in this great episode (see Fig. 7).
Invasion occurred in both directions. By a moderate tabulation, fifteen families of North American mammals then spread into South America and seven families spread in the reverse direction. The main migrants to the south were rabbits, squirrels, field mice, dogs, bears, raccoons, weasels, cats, mastodons, horses, tapirs, peccaries, camels, and deer, including in most of these cases some variety of related forms.
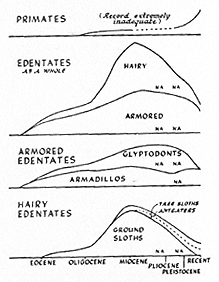
Fig. 8. Development of some South American groups that were ecologically unique with respect to late invaders from North America. Height of the graph in each case represents the relative number of known genera. The time scale at the bottom applies to all graphs. NA = migration to North America. (Some of the old native rodents were also ecologically unique; see Fig. 9.)
The immediate effect was to produce in both continents, but particularly in South America, a greatly enriched fauna. To a fauna already large and essentially complete or closed ecologically were added a large number of new forms from the other continent. The enrichment inevitably involved some duplication. No two forms of different origin can have been precisely and fully equal in their needs and capacities, but many were sufficiently similar to be in competition for food and, in general, living space. Some native groups held their own and some invading groups became extinct, but some native groups disappeared and (as a rule) were replaced by invaders.
In the end, that is, at the present time, South America has returned to about the same basic richness of fauna as before the invasion. Recent families of mammals there number the same (more or less, depending on the classification used) as in the Pliocene before invasion, but the faunal composition is radically different. The notoungulates, litopterns, and marsupial carnivores have entirely disappeared. The native rodents and edentates are greatly reduced. In their places, artiodactyls, perissodactyls, rodents of northern origin (squirrels, cricetids), rabbits, and placental carnivores are fully entrenched and constitute, in number of genera, about half of the recent fauna.
[[p. 382]] The main determinants in this process were, first, ecological status and, second, place of origin. Ecological uniques tended to survive (Fig. 8). It is true that glyptodonts and ground sloths, after spreading over both continents, became extinct and that they apparently were ecological uniques. The question of their extinction involves some other and not properly understood factor, and it is part of a larger question that cannot be discussed here. The smaller, likewise or even more strictly ecologically unique relatives of these animals, armadillos, tree sloths, and anteaters, did survive. So did the monkeys, ecological uniques with respect to North America, and most of the truly ecologically unique old native rodents.
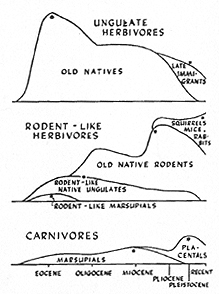
Fig. 9. Development in South America of ecological vicars, similar in adaptation to invading types from North America. Height of the graph in each case represents the relative number of known genera. The time scale at the bottom applies to all graphs. Top and bottom graphs show simple, total replacement of old groups by late invaders from North America. The more complex middle graph shows total replacement of old rodent-like marsupials (Polydolopidae) by other old native rodent-like forms, partial replacement of old rodent-like ungulates (mainly typotheres and hegetotheres) by old native true rodents, and then partial replacement of most of the latter by rabbits and true rodents in the late invasion from North America. Surviving old native rodents are, in the main, ecologically unique, with no closely similar competitors among the late invaders. * = apogee.
When ecological vicars met, one or the other generally became extinct (Fig. 9). In South America, old native ungulates disappeared and ungulates of northern origin survived. Many old native rodents (about half the generic lines) became extinct and rabbits, squirrels, and field mice survived. Marsupial carnivores became extinct and placental carnivores survived. The fact that in each case the survivors were of northern origin cannot be pure coincidence. It is not explanatory to say that the animals from North America were "superior" or "more progressive," and such statements would be hard to substantiate by any objective evidence from their anatomy, for instance.
The ultimate factors have not been and probably cannot be designated, but a generalized explanation presents itself. North American animals had intermittently throughout the Age of Mammals and almost continuously in its later part been involved in the flux and intermigration of the World Continent. Those extant in the Plio-Pleistocene were the ones that had been successful in a long series of competitive episodes. They were specialists in invasion and in meeting competitive [[p. 383]] invaders. South American mammals had competed among themselves in the early Tertiary, but by about the end of the Oligocene they had essentially completed a process of parceling out the ecological opportunities among a number of practically noncompeting lines. Thereafter until the late Pliocene they met no impact from outside their own closed economy, and when it came, they had not evolved the required defenses.
Thus the recent South American mammalian fauna is a complex agglomeration, in spite of the fact that processes of ecological adjustment have again reduced it to an essentially balanced economy. It may all be ultimately derived from North America, and some of the evidence to that effect has been summarized. This evidence is suggestive but inconclusive for the oldest elements, stronger for the mid-Cenozoic elements, and conclusive and unquestioned for the latest elements. Even if all did have the same geographic origin, there are the three quite distinct broad faunal strata. Their different times of entry into South America complicate the picture, not only because of corresponding differences in differentiation within that continent, but also because each stratum was drawn from a different sort of World Continent fauna. Differentiation, replacement, intermigration, complex stratification, divergent specialization, and other processes were constantly going on in the World Continent, and the strata of South American mammals sampled this sequence at three very different levels.
The static zoogeographic picture in a classical sense, of current resemblances and differences of regional faunas, is further complicated by differential regional extinction. Pumas link North and South America in nonhistorical zoogeography because they now occur on both continents, and camels separate the faunal regions, because they occur in South but not in North America. But in the Pleistocene pumas and camels occurred throughout both continents and in the Pliocene their ancestors were present in North but not in South America. Final analysis of the present relationships of South and North American faunas must involve this factor as well as the factor of stratification.
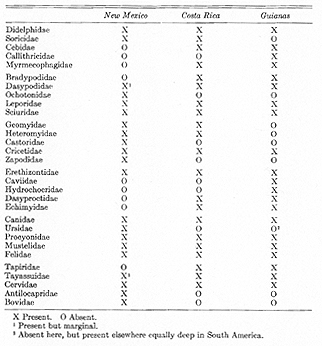
In Table II an attempt is made to analyze the now extant South American mammals with respect to their geographic history and their present relationships to mammals in the recent North American fauna (north of the tropical zone).
This analysis is on a family level. More refined analysis, to genera or below, would involve some changes of status in the last stratum. Central America, with its peculiar faunal history and its intermediate faunal types, is not involved in this comparison, which is based on the fully South American and the temperate-zone North American faunas. Special consideration of Central America is now necessary.
The Role of Central America
There is and doubtless always has been considerable difference between regional faunas within South America. The faunas of the Patagonian pampas, the Andine punas, and the tropical rain forests are strikingly [[p. 384]] distinct. They are, nevertheless, regional varieties or differentiates of a general South American fauna and they share much the same sort of differences from and resemblances to the general fauna of temperate North America. There have not, during the Cenozoic, been any absolute barriers between regions within South America, and no outstanding effects of isolation are historically evident in separate parts of the
TABLE II--ANALYSIS OF ZOOGEOGRAPHIC RELATIONSHIPS OF SOUTH
AMERICAN RECENT MAMMALS, WITH RESPECT TO NORTH AMERICA
I. Oldest South American faunal stratum, from North America
(?) in late Cretaceous-Paleocene.
A. Collaterals (strongly differentiated)
from same level present in North America.
1.
Without late level spread and recent presence in North America: caenolestids
(North American Colaterals: opossums). (Some students would place the
caenolestids under IB1 and the opossums under IB2; this may be correct
but seems unlikely at present.)
2.
With late level spread, etc.: none.
B. Ancient stock extinct in North
America.
1.
Without late level spread and recent presence in North America: none.
2.
With late level spread, etc.: armadillos (N. A. distribution restricted).
II. Intermediate South American faunal stratum, from North America in
Oligocene.
A. Collaterals (well differentiated)
from same level present in N. A.
1.
Without late level spread, etc.: "hystricomorphs" or caviomorphs except
porcupines (differentiated N. A. collaterals: sewellel, squirrels, etc.).
2.
With late level spread, etc.: porcupines.
B. Ancient stock extinct in North
America.
1.
Without late level spread, etc.: monkeys. (No collaterals in N. A. wild
fauna, but man could be considered as such.)
2.
With late level spread, etc.: none.
III. Late South American faunal stratum, from North America in late Pliocene
and Quaternary.
A. Collaterals from same level (here
poorly differentiated and representing essentially the same immediate
stock) present in N. A. Late level spread back to N. A. possible in some
cases, but in that event part of the same general episode of intermigration
as emplacement of the South American stratum: opossums (history somewhat
questionable, sometimes considered IB2, but probably belonging here),
shrews (marginal and with slight penetration in S. A.), rabbits, squirrels,
cricetids, dogs, bears, raccoons, weasels, cats, peccaries (marginal and
with slight penetration in N. A.), deer.
B. Collaterals extinct in N. A.: tapirs,
camels. (Peccaries, listed under A, are marginal between A and B.)
continent. At most there has been a sort of climatic zoning by which animals once more widespread have become confined to particular, areally definable environments (primates to tropical forests, camels to mountains and cold plains, etc.), or the faunas of deteriorating environments have been progressively impoverished (in Patagonia, for example). Mid-Cenozoic faunas lately found in Colombia (cooperatively by the [[p. 385]] Colombian government and the University of California, under the direction of R. A. Stirton) have interesting regional differences from contemporaneous Patagonian faunas, but are definitely of the same general type and reveal no unexpectedly exotic groups. An old idea that northern and southern South America were separated by a Tertiary sea barrier and had quite distinct faunal histories, an idea still current among a few students but long rejected by others, is thus conclusively proved false.
There are, nevertheless, two regions of (broadly speaking) Latin America and of the Neotropical Region of static zoogeography that have had faunal histories decidedly different from that of South America: Central, or in a somewhat broader sense Middle, America and the West Indies. The development of the West Indian fauna is one of the most fascinating topics of historical zoogeography and has been the subject of long, sometimes bitterly polemic discussion involving fundamental principles of this science. It is, however, of minor importance for the present broader theme and cannot be considered here. The West Indies have been a faunal dead-end. There is little evidence that they have had a reciprocal influence on the larger faunas of the continental mainlands.
Middle America, on the contrary, has had an essential and striking role in the development of the faunas of all the Americas. A few admirable studies of the static, modern zoogeography of parts of the region have been made, and some attention has been given to historical aspects, but full evaluation is lacking and too little thought has been given to this subject. A cause for relative neglect has been the extreme paucity of primary historical documents. With one possible exception of no present value, no pre-Pliocene nonaquatic fossil has ever been found in tropical Middle America. There is one known, rather small but extremely important, early Pliocene mammalian local fauna, from Honduras (found and described by P. O. McGrew and associates), and there are several scattered Pleistocene faunules, of considerable interest but still lacking forms (especially the smaller mammals) that would be more fully enlightening.
Direct and conclusive study will require a good sequence of Central American land faunas, including vertebrate microfaunas, from Miocene, at latest, to recent. There is no assurance, nor even any considerable probability, that such a sequence remains to be discovered. It is nevertheless possible to evaluate the role of this region with some assurance, even if without adequate detail, on the basis of the scraps of local, direct evidence and the increasingly imposing array of indirect evidence.
The recent fauna of Central America is essentially like that of South America. The same faunal strata occur there, although, as already noted, their ages and relationships with respect to the area occupied are here different. Environmental conditions in Central America are similar to those of adjacent tropical South America. With union of these two areas, the resulting neotropical fauna has occupied territory as far as the environmental similarity extends. This is roughly as far as the climate can be called "tropical" in broad terms, and the line [[p. 386]] conventionally bounding the Neotropical Region to the north has been drawn at the equally conventional boundary of tropical climates, delimiting an area extending to and stopping with the hot Mexican lowlands. This is, indeed, merely a critical line within a broad transitional zone and not a localized barrier. Few distributions stop precisely at the line. Starting far north of it, say in Colorado or New Mexico, and
TABLE III--FAMILIES OF RECENT LAND MAMMALS IN
SELECTED PARTS OF NORTH, CENTRAL, AND SOUTH AMERICA
progressing to regions far south of it, say to Ecuador or the Guianas, animals mainly neotropical in distribution rather steadily become more frequent, and those mainly nearctic less frequent. In detail, Central America also has local differentiation, with numerous species and some genera (but no higher categories among mammals and few among other animals) confined to that area.
The place of Central America within the broad outlines of static recent [[p. 387]] zoogeography is well displayed by comparison of the mammalian families of New Mexico, Costa Rica, and the Guianas, each area with the same number of families (twenty-one). About half the families (ten fully and two more marginally) are common to all three areas but generally with different species and sometimes with different genera in each. Six families occur in New Mexico only, as among these three areas, and three in the Guianas only. These are the most definitely nearctic and neotropical groups, respectively. Costa Rica has no families that do not occur either in New Mexico or in the Guianas. It has three families in common with New Mexico but not the Guianas and six with the Guianas but not New Mexico, and is thus intermediate in this respect but somewhat more like the Guianas, justifying inclusion in the Neotropical Region (see Table III).
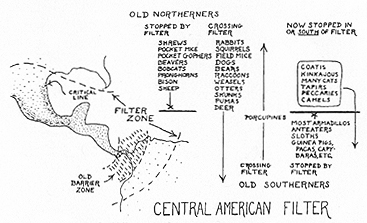
Fig. 10. Diagram of middle America as a faunal filter. The examples given are from the recent fauna of land mammals. For some groups the action of the filter was different in the Pleistocene.
Middle America may be considered statically as a transition zone and historically as, successively or intermittently, a barrier and a migration route, but these simple and usual characterizations hardly begin to express its true role and importance.
As a transition zone and migration route, the role is not merely that of a habitat and a pathway. Middle America is a faunal filter (see Fig. 10). Its ecological characteristics, in the broadest sense, determined which stocks were involved in faunal interchanges between North and South America and which are now immobilized to north and to south. The filtering action is not sharply localized. It begins well to the north (and west), roughly at the edge of the lower Sonoran life zone in southwest United States, and also reaches far to the south and east, more or less to the edge of the Guiana highlands and thence southward and westward. From these quite indefinite outer edges, the filtering action becomes denser toward the central critical line, which now [[p. 388]] approximates the border of the central Mexican plateau. It probably had about the same or at times a more southerly position in the Pleistocene, but in parts of the Tertiary it may have been considerably farther north.
At present some northern groups, such as the shrews, pocket mice, and pocket gophers, penetrate well into the filter zone, but not beyond its vague southeastern edge. Others, such as the bobcats, bison, and sheep, have stopped near its attenuated northern border. For other northern forms, the filter zone has been essentially an open passage: rabbits, squirrels, dogs, otters, pumas, and many others among recent animals, and also horses, mastodons, and some other fairly recently extinct groups. Most of the old southerners, South American mammals of the first and second faunal strata, are now stopped somewhere within the zone: most armadillos, all anteaters and tree sloths, all the old native South American rodents except the porcupines. One armadillo (Dasypus novemcinctus) is peculiar in that a generation or so ago it stopped at about the northern edge of the filter zone but in the last few years has spread well beyond this. A related form (Dasypus bellus, now extinct) was even more widespread in North America in the Pleistocene, when there were also a few other old southerners (glyptodonts, ground sloths, capybaras) unaffected by the filter. It is also interesting that the filter was in some cases permeable for ancestral forms but now completely separates their differentiated northern and southern descendants, for instance in the cases of the porcupines and the bears.
Most peculiar in this respect are the rather numerous animals clearly of ultimate northern origin and yet now stopped in or south of the filter zone: coatis, kinkajous, the numerous kinds of South American cats other than the puma, short-faced bears, tapirs, peccaries (reaching the extreme northern edge of the filter, but hardly beyond), camels, and other lesser differentiates. Superficially one can say simply that such groups happened to become extinct in North but not in South America, a statement but not an explanation. For most of them, at least, there is a more explanatory probability: the particular populations and lines involved were native to Middle America, adapted to environmental conditions prevailing there and over much of South America and not adapted to or immediately derived from North America above the present filter zone. In some cases there are complications requiring modifications of or additions to this general theory (for instance, for the bears, paramo tapirs, and camels), but even these need not be flatly exceptional or contradictory, and for most groups involved the theory is a simple, elegant, and sufficient explanation of their peculiar distributional history.
It is an obvious, but nevertheless frequently unappreciated, fact that immigrants from North to South America did not come from the continent as a whole or from its broad, now temperate zone, best known to us both paleontologically and neontologically, but only from Central America. With local and geologically brief interruptions for part of it, Central America has been continuous with the North American land mass throughout the Cenozoic. Marine barriers were mostly in the [[p. 389]] extreme southeastern part of Central America and the major barrier, especially toward the end of the Cenozoic, was in what is now part of South, not Central, America (in western Colombia). Thus Central or broadly Middle America must have had a fauna mainly or purely North American in origin and in broad faunal type until toward the end of the Pliocene, a conclusion usefully attested by the one known Tertiary land fauna from Central America, early Pliocene in age and entirely North American in affinities. Nevertheless, Middle America must have been an important center of regional faunal differentiation within the North American general fauna.
Even in the early Cenozoic, when climatic zoning was less sharp than now, such zoning did exist, and Middle America has by astronomical and meteorological necessity always been the warmest (or most evenly warm) and tropical part of North America. It must long have been a center of adaptation and local radiation of faunal elements specifically adapted to its special conditions, not only climatic but also edaphic, floral, etc. This special local fauna was the one, and the only one, available for spread to South America, in all of the faunal strata of the latter region. Some Central American differentiates have succeeded in spreading northward (jaguars in the Pleistocene, for instance, and probably a number of otherwise mysterious newcomers in more northern fossil fields throughout the Cenozoic). Spread southward, when geographically possible, was more general and rapid because northern South America is more like Middle America in climate and associated factors. The Tertiary sea barrier did not follow a climatic zonal boundary, and when connection was established the incorporation of Central America into the South American and of South America into the Central American faunal zone was rapid and reciprocal.
Thus may be readily explained neotropical differentiation within many groups of old northern origin, the roots of which have not been found among the fossils from higher latitudes in North America. The more distinctively neotropical forms (at levels mainly of specific to generic differentiation) in many families may be inferred to have been Middle American differentiates: numerous noncaviomorph rodents, some dogs, the southern procyonids (sensu lato), most of the southern cats and most of the deer, among others. There is also discernible, on these grounds, a minor stratification within the broad late stratum of the South American fauna, between forms which had been longer in Middle America and more differentiated there, and those newer or less isolated in that region. Thus the coatis (Nasua) are inferred to be older and their allies the true raccoons (Procyon) younger Middle American forms, or among the deer the brockets (Mazama) may be older and the closer allies of our white-tails (Odocoileus) younger there. In some such cases there are other factors to consider, especially the possibility and timing of spread northward from Middle America. Here emphasis can only be placed on broader aspects of the historical role of Middle America and details cannot further be discussed. In fact, these details have not yet been adequately studied, if at all, from this point of view.
*
*
*
*
*