
http://people.wku.edu/charles.smith/biogeog/SIMP1936.htm
Data on the Relationships of Local
and
Continental Mammalian Faunas
by George Gaylord Simpson (1936)
Editor Charles H. Smith's Note: Original pagination
indicated within double brackets. My thanks to The Paleontological Society
for permitting the reprint. Citation: Journal of Paleontology 10
(1936): 410-414.
[[p. 410]] Abstract
A comparison of Recent mammalian faunas of Florida and New Mexico, two remote regions differing essentially in climate and topography, shows that knowledge of either fauna will give a fair idea of the general composition of the other. It seems probable, for example, that such a fauna as that of the Bridger formation gives an excellent representation of the fauna of North America as a whole in medial Eocene time, and that broad inferences drawn from it are of higher value than is generally granted.
The notorious incompleteness of the paleontological record is due not only to gaps in time distribution of known fossils, but also, and at present in still greater degree, to the relatively local character of most of the faunas known. It is often necessary to judge the mammalian fauna of a whole continent or major land area from fossils found only at one or a few localities. For instance, the Eocene land life of North America is known almost exclusively from discoveries in a few basins scattered through the Rocky Mountain area. It is usual to speak of the Eocene fauna of North America, but in fact for any one part of the Eocene only a relatively very small and localized geographic group of that general fauna is known. Other instances are even more striking, as in South America, where almost all known pre-Pliocene mammals are from the relatively limited and marginal areas of Patagonia. It is obvious that such data cannot give a complete picture of the fauna of a whole landmass.
In the endeavor to visualize these relationships and to give some concrete basis for their evaluation, detailed comparison has been made between the Recent mammalian faunas of Florida and of New Mexico. The results, here set forth in outline, clearly are no more than an approach to the general problem, which requires very many more data for any full consideration or definitive solution. They do, however, suggest a means of approach which does not appear to have been utilized in this way previously, and they give some definite basis, however inadequate as yet, for judgment in an important matter hitherto generally ignored or treated by guesswork.
Florida has a land area of 54,861 square miles and a water area of 3,805 square miles. It is a marginal region and is almost surrounded by the sea. Physically it is low, with little relief or physiographic variety (maximum elevation, 325 feet; minimum, sea level; average 100 feet). The climate is oceanic and is warm and humid. Faunally, parts of the Lower Austral and Tropical zones [[p. 411]] are included. The mainland lies between 25°07' and 31° north latitude.
New Mexico has a land area of 122,503 square miles and a water area of 131 square miles. It is central in position and far from the continental borders. The physiography is extremely varied, including high mountains, plateaus, plains, desert basins, and low alluvial valley bottoms (maximum elevation, 13,306 feet; minimum, 2,876 feet; average, 5,700 feet). The climate is markedly continental, with hot summers and cold winters, and in general is semi-arid. It includes parts or representatives of all fauna zones from Hudsonian to Lower Sonoran. It lies between 31°20' and 37° north latitude.
The closest points of the two states are over 900 miles apart, and their centers are separated by about 1,450 miles. They are markedly different in position, environmental and physiographic variety, climate, and faunal zones. It is probably impossible to duplicate exactly in either an environment found in the other. They have in common the facts that both are distinctly parts of the North American continent of long standing, from the point of view of recent mammals, and that both lie in the southern part of the Temperate Zone.
The primary object of the comparison is to see what could safely be inferred of the fauna of the other if the mammalian fauna of only one of these two states were known. Hence some judgment is possible as to the limits of safe inference regarding the regional fauna in cases of fossil faunas known only from one part of a large landmass.
Lists of recent groups of land mammals of the two states have been compiled, and while no two students would agree as to exact numbers, the relative proportions are not greatly dependent on any personal factor. The following figures represent the number of orders, families, genera, and species (subspecies being excluded) of recent terrestrial mammals, exclusive of man and his importations, native to each state, and the numbers common to both.
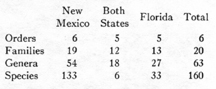
The data for each state are more graphically comparable in fig. 1. The immediately striking fact is the greater variety of the New Mexican fauna, with four times as many species as Florida. The apparent reasons for this, in probable order of importance, are the greater environmental diversity of New Mexico, its larger area, and its more central position.
A second noteworthy fact is the markedly different ratios for different taxonomic groups. New Mexico has four times as many species as Florida, but only twice as many genera and half again as many families, while the number of orders is nearly the same. The two curves of the diagram consequently converge to the right. If comparable curves are plotted for areas still larger or still more varied than New Mexico, they
[[p. 412]]
rise more and more steeply to the left, but the convergence to the right remains.
As regards this example, then, if we knew all the mammals of one state, we could infer almost exactly how many orders occur in the other, and fairly well how many families, but might be badly mistaken regarding the lesser units.
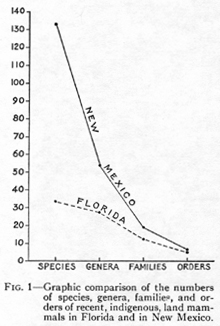
More important than the number of units is their character. Somewhat contrary to expectations, the example suggests that in similar cases and within certain broad limits, the character of the fauna of one region can be inferred from that of the other. These data are graphically shown in fig. 2, in which the elements common to both states are contrasted with those peculiar to one. Another form of presentation is that of this table:

On this showing, knowledge of either fauna will give a fair idea of the general composition of the other, that is, of its orders and families. The possibility is also enhanced beyond the point indicated by the figures, and especially as regards smaller groups, by the fact that many mammals of each fauna have relatives in
[[p. 413]] the other which are not identical but which are so similar that knowledge of one is applicable in a general way to the other.
Another striking fact, although one that might be assumed a priori, is that the larger fauna gives a better basis for inference as regards the smaller than does the smaller for the larger. New Mexico, with its highly varied fauna, gives a reasonably accurate picture of the mammalian life of at least the central part of North America. If we knew its fauna and not that of Florida, we would nevertheless be familiar with all the orders, almost all the families, and two-thirds of the genera that do occur in Florida. Even beyond this, although for instance, we would not know Neofiber or Geomys, we would have the closely related and ecologically similar genera Ondatra and Thomomys. There would in fact be only one Floridian genus, Scalopus, which was radically unlike any known from New Mexico.
Applied to paleontological data, the accuracy of such inferences depends on numerous factors which can be approximately evaluated, although any close approach to statistical accuracy would necessitate the accumulation and interpretation of many more data than have yet been studied, and would even then be an extremely complex and difficult problem. There is, however, no reason to believe it insoluble. The main factors requiring such consideration are probably the following:
1. The area represented by the collection. This can be calculated, at least approximately, by the actual area covered by the geological formation, or by the probable size of the drainage area from which its fossils could have been derived. Accurate evaluation of this factor demands statistical data on the relationship of area and number of species under similar recent conditions.
2. Environmental variety. This can be estimated from the inferred physiographic conditions when the formation was being deposited, or perhaps more accurately from the nature of the fauna. A fauna which includes many cases of related but truely distinct contemporaneous species surely indicates environmental variety, for in general only one such species can live in one local environment, in one region, at one time (Cabrera's law of "ecological incompatibility"). Furthermore paleobiological inference may also indicate environmental variety, by indicating variety, or lack of variety, in the probable habits of the animals found.
3. Geographic position. It can be reasonably established on paleogeographic grounds and sometimes in part from the fauna itself, whether the region from which a given fossil fauna is derived was at the time of that fauna part of the large landmass under consideration, and approximately what position it occupied on that landmass. Presumably the more central the area, the more varied its fauna is likely to be and the better it will represent the continental fauna as a whole. The influence of broad climatic zones also needs evaluation. Presumably a large fauna would include nearly 100 per cent of the important groups of mammals present on the same continent in the same climatic zone, and in passing to other climatic zones the percentage would be progressively lower. Within a reasonable margin of error, this relationship could probably be established mathematically. In the case of Tertiary faunas the factor would be less important than with recent faunas, for the climatic zones were evidently much less marked during most of the Tertiary than they are at present.
[[p. 414]] 4. Completeness of single faunas. Accuracy of broad inferences must be directly proportional to the extent of knowledge of the fauna on which the inference is based. It is probably impossible in any case to obtain fossil representatives of 100 per cent of the groups of land mammals that actually lived in a given area. The percentage of representation can be roughly estimated from the size of the collection made, the percentage of new species added by new collections, the relation of the number of species known to the number which probably were present under the given conditions, the presence or absence of representation of given groups known to have occurred both before and afterwards in the same region, and the ecological character of the fauna, that is, whether all environments probably present are represented, whether both herbivores and carnivores are present in normal proportions, whether both large and small mammals are present, etc.
It seems probable that a large and varied Tertiary fauna which may be judged representative on these criteria, even if it is from a single and fairly local geological formation, gives an excellent representation of the mammalian orders present on the whole continent, a good idea of the families, and will probably even include most of the genera.
As an example, the mammals of the Bridger fauna probably ranged over an area comparable to that of Florida. The position of their area was very central on the Eocene continent. The local environments represented are clearly numerous. Knowledge of the fauna is fairly complete. It follows that this fauna probably well represents the middle Eocene life of all of North America.
As an example of the sort of broader and more important inferences dependent on such data, the well known and apparently representative middle Paleocene faunas contain no possible ancestors of several Eocene orders and lesser groups. From the considerations here given, it therefore seems highly improbable that many of the typically Eocene groups were then present in North America, and failure to find them in the Paleocene cannot be entirely imputed to the localized nature of the known deposits of middle Paleocene age.
It seems probable that the paleontological data now in hand provide a better basis for studies of mammalian faunal succession and geographic distribution than is generally granted.
*
*
*
*
*