
http://people.wku.edu/charles.smith/biogeog/PILS1901.htm
The Genesis of Mid-Pacific Faunas
by Henry A. Pilsbry (1901)
Editor Charles H. Smith's Note: Original pagination
indicated within double brackets. Notes are numbered sequentially and
grouped at the end, with the page(s) they originally appeared at the bottom
of given within double brackets. Citation: Proceedings of the Academy
of Natural Sciences of Philadelphia 1900 (Vol. 52): 568-581.
[[p. 568]] The new
system and implied phylogeny of the Vasopulmonata outlined in my paper
on Partula and Achatinella throw a new light upon the
constitution of the island faunas of the Pacific, and reopen the whole
discussion of their origin and distribution.
Within the last decade a South Pacific continent, stretching its great arc from New Zealand to Chili, has been postulated by Dr. von Ihering1 and Professor Hutton,2 while Dr. G. Baur3 has also adduced evidences toward a similar conclusion. This hypothesis will be alluded to below. The views of Wallace regarding these faunas, based upon the absence of amphibia and terrestrial mammals, and upon certain geological considerations, have been accepted by the great majority of zoogeographers, who hold that the Polynesian faunas have been forever insular, and have derived their constituents from circum-Pacific continents by over-sea drift, and by wind or birds carrying animals or their eggs.
To the student of vertebrates such conclusions seem justified by the facts; but a little reflection shows one that opinions based upon the distribution of vertebrates may require radical revision when [[p. 569]] invertebrate groups are taken into consideration. This is especially true in zoogeographic matters because the higher groups have changed rapidly, the lower slowly. Many genera of land snails reach back to the oligocene unchanged save in specific characters, and there can be no reasonable doubt that the modern family groups of these snails diverged far back in mesozoic time; yet how complete a revolution the brief pliocene wrought in mammalian life! A land mass isolated in, say, mid-tertiary or oligocene times, would necessarily be lacking in many important mammalian genera and families developed subsequently, yet it might be quite identical in molluscan genera with the tract from which it was insulated. The existing families and genera of even the reptiles and batrachians do not have the antiquity of groups of the same nominal rank among non-marine mollusks. But no especial argument need be made here in support of the thesis that the rate of structural differentiation has been constantly accelerated as animals rose higher in the scale; and therefore it follows that zoogeographic "provinces" based upon the distribution of land snails or earthworms, for instance, recall older arrangements of sea and land than those based upon the distribution of terrestrial vertebrates--mammals and birds.
Now upon glancing at the table of mid-Pacific faunas on p. 576, the student of continental molluscan faunas will first notice the great rarity of Holopoda, and the total absence of Agnathomorpha and Agnatha, and of the more specialized and presumably late Aulacopod families Limacidcæ, Arionidæ, Philomycidæ, as well as the highly organized forms of Zonitidæ with complicated genitalia, such as the Ariophantinæ. This leaves a fauna composed almost exclusively of Orthurethra, Elasmognatha, and the less specialized families of Aulacopoda. The land operculates are chiefly Realiidæ and Helicinidæ; and in the fresh-water faunas, Limnæa, Planorbis and Physa of the continents give place to "Bulinus," a stock probably ancestral to Planorbis; and "Melania" alone represents the prosobranchs.
The significance of a fauna so constituted becomes startling when we consider that the Orthurethra are a little-changed remnant of the parent stock of all Vasopulmonata; the Elasmognatha are an early branch of this stock, while the Aulacopoda are unquestionably, on anatomical grounds, the lowest branch of the Sigmurethra [[p. 570]] or ordinary land snails.4 Is not the whole fauna what we would expect in a region populated near the time of divergence of the earlier groups of Vasopulmonates, and isolated since? Or, to state the question in its bald entirety:
Are the mid-Pacific snail faunas witnesses to the existence of a palæozoic or early mesozoic land mass, probably continental in proportions, and peopled by representatives of nearly all land-snail groups then existing?
A negative answer implies the alternate hypothesis that these faunas are assemblages of waifs, the survivors of many a precarious voyage. An affirmative answer should give reasons for the supposition of more extended land area than now exists, and for the geologic date assigned.
The primitive or antique character of the Polynesian faunas, commented on above, may be graphically represented by a table, in which the families and genera of land Pulmonates of continents
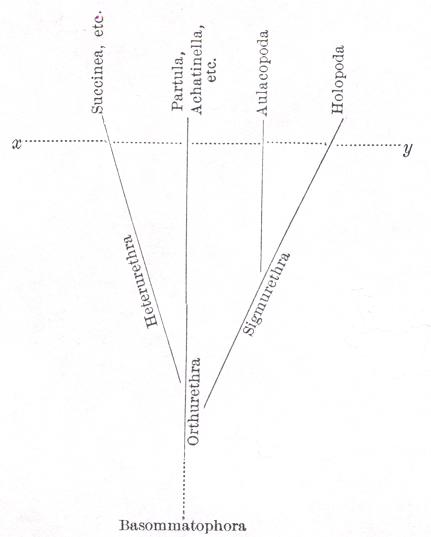
The line x-y may represent the degree of differentiation reached when the Pacific land-faunas were established. Subsequent differentiation is not represented on the diagram.
[[p. 571]] or continental islands are contrasted with those of the mid-Pacific. I select continental North America, Great Britain and the Philippine Islands for this purpose, merely because good recent lists lie on my desk;5 but practically the same results would be seen were I to contrast South America, the Antilles, Africa, Europe or Australia with the Pacific archipelagos.
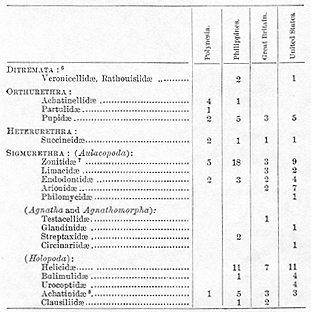
[[p. 572]] Note that the Sigmurethra in North America number 11 family groups and 47 genera (with a vast preponderance of species), to 3 families and 7 genera of all other (lower) Stylommatophores. In Polynesia there are only about as many genera of Sigmurethra as of Orthurethra, and nearly the whole of the former belong to low and primitive types of Aulacopoda.
But, it may be objected, oceanic faunas are thus unsymmetrical from the very nature of the accidental means by which oceanic islands have been populated. Granted; but is there any reason why the occasional and unusual vehicles by which such population is effected should act with exclusive discrimination against whole series of higher groups, in favor of the lower? It is very easy to show that snails may have been carried from place to place by a hurricane, a floating tree or "floating island,"or their eggs may find room in the pellet of earth clinging to a bird's feather; but it is incumbent upon the theorist who peoples the mid-Pacific islands by such means to show why such dominant groups as the Helicidæ, Bulimulidæ, Rhytididæ, Streptaxidæ--in fact, the whole Holopoda and Agnathomorpha, with higher members of Aulacopod families, as well as the higher operculates--should have utterly failed to take advantage of these means of transport.
To the possible objection that low forms survive an ocean voyage better than high, may be urged the fact that such has not been the case in Madeira, the Azores, Cape Verdes, St. Helena or Bermuda, in all of which Holopod groups occur, and in many cases were established long enough ago to become generically or specifically differentiated. To the further objection that the primitive groups may adapt themselves more readily to new environments, the same argument applies. Moreover, it is well known that the most successful emigrants of all snails are species of the highly organized or specialized Helicidæ, Achatinidæ, Limacidæ and Arionidæ, which have followed modern agriculture and commerce all over the world. Although a number of snails have been introduced by commerce into Polynesia from the East Indies, Europe or America, I know of no single case of a Polynesian snail becoming acclimated in any other part of the world.
The advocate of a Polynesian waif fauna is then compelled to adopt the view either that accessions to the mid-Pacific snail faunas practically stopped a long time ago, from causes unknown or [[p. 573]] hypothetical, or that an unparalleled series of accidents intervened to prevent the, in recent times, ubiquitous and prominent continental groups of snails from effecting a landing in these islands.
Wallace derives the Polynesian faunas from his Australian region, mainly on the evidence of the birds. But no such idea can be entertained regarding the snails. In a rich development of Endodontidæ, the two areas agree; but that family is of worldwide distribution and known great antiquity, and therefore proves nothing. Nowhere in Australia, New Zealand or adjacent islands do we find a parent stock for the most characteristic of Polynesian groups, the Orthurethra. Australia itself has of this group only modern continental types of Pupidæ--Pupa, Pupoides and Bifidaria,9 which probably reached that continent with the epiphallogonous Helices, from an East Asiatic center. If there were earlier Australian Orthurethra, they became wholly extinct before the higher snail groups which have long occupied Australian soil. Most of the Pupidæ (except Bifidaria) and the few other Orthurethra, such as Tornatellina and Partula, which extend into Melanesia, etc., are Polynesian in their affinities, and evidently to be regarded as outlying colonies from that centre, derived from the western edge of the Pacific land. Far from being a faunal dependency of the Australian or Oriental regions, Polynesia has every appearance of being a region which started with a fauna long antedating the present Australian and Oriental faunas, developing along its own lines, retaining old types because they did not come into competition with the higher groups developed on the greater and less isolated continents.10 It is significant that the typical Polynesian groups attain their acme of structural as well as specific differentiation in the mid-Pacific Society and Hawaiian groups. Were these faunas derived from waifs drifted, blown or otherwise carried from the Australian or Oriental regions, we might reasonably expect a diminution from the West outward, such as the Azores show compared with Madeira, or Bermuda compared with the Bahamas.
On the supposition that Polynesia has always had the [[p. 574]] constitution the name implies, it is difficult to see why agencies which introduced representatives of some eight families of snails into the Hawaiian group should totally fail to act during the late mesozoic and tertiary. Even Wallace felt that some explanation was called for, and speaks vaguely of the "extensive shoals to the south and southwest" and "two deep submarine banks in the north Pacific between the Sandwich Islands and San Francisco."11
Dr. G. Baur, in discussing the Galapagos fauna, applied the terms "harmonic" and "disharmonic" to island faunas and floras. The fauna or flora of a continental island will be harmonic, because it will share the several groups of the parent continent in due proportion. An island elevated from the sea bottom and not connected with other land will have a disharmonic fauna and flora--"that is to say, it will be composed of a mixture of forms which have been introduced accidentally from other places."
It is almost inconceivable that a number of widely separated islands of oceanic origin should agree or be harmonic among each other in their faunal constituents; but taking the Polynesian groups included in or adjacent to a triangle with the Hawaiian Islands, Marquesas and Samoan groups at the angles, we find a remarkable agreement between their faunas. Comparing any one with the nearest continental faunas of to-day, they seem disharmonic; but among themselves there exists a remarkable homogeneity; and if we compare them with what the continental faunas must have been shortly after the divergence of the Holopod from the Aulacopod Sigmurethra, it will be seen that Polynesia now possesses representatives of what must have then been all the main groups of Vasopulmonata; and the preponderance of Orthurethra is precisely what would be expected.
The harmonic characters of these faunas is the more striking when we compare them with the Atlantic islands admitted to be "oceanic"--Madeira, the Azores and the Cape Verdes. In these islands the genera are all clearly derived from Eur-African sources, being all either genera still existing on the mainland, or more nearly allied to existing or tertiary European groups than to those [[p. 575]] of any other part of the world. Only two genera are common to the three island groups and the mainland, and but one other genus is common to the three archipelagos. Madeira and the Azores have 7 genera in common, and Madeira and the Cape Verdes 7; only 4 of these being identical in both cases, and this in a total fauna of 21 genera, 16 of which are common to Europe and one or another of the island groups. We eliminate from the account all known or presumed introductions during the period of commerce. The sporadic or disharmonic character of the faunas is strikingly illustrated by the almost omnipresent Palæarctic genera Clausilia, found only in Madeira; Helicella, wanting in the Azores, and, further, by the distribution of Cochlicopa, Helix, Punctum, and especially Succinea and Buliminus, both groups of great antiquity, the former found on almost all islands, but wanting in the Azores and Madeira. The presence of slugs (Plutonia) on the outlying Azores, and their absence in Madeira (until Limax was introduced by commerce) is also notable.
The distances are much less than those sundering many of the Pacific archipelagos, and yet their faunas are less harmonic than those of Polynesia. And still in all probability the older elements of these Atlantic waif faunas do not much precede the tertiary, if at all, and additions by natural causes have continued to, I suppose, the present time. Indeed, part of the common element in the three island groups may fairly be traced to a common ancestry in the European tertiary, rather than to actual communication between the archipelagos themselves.
[[p. 576]] Contrast, now, a table of seven widely separated Polynesian faunas:

The remarkable homogeneity of faunas scattered over so wide an area as Polynesia, indicates either a former great extension and therefore approximation of the archipelagos, or that they actually formed parts of a single land mass. Although the distances between them are generally far greater than separate the Atlantic island groups, the faunas show a much greater common element; and had they been derived from similar sources it is difficult to see why the Atlantic faunas should be so much less harmonic than the Pacific.
The former moderate extension of the Hawaiian and other islands considered probable by Wallace, while a step in the right direction, is insufficient to account for the facts. The faunas are, in [[p. 577]] fact, what might be expected were they remnants of a sunken continent; and the waif theory signally fails to account for their composition, although no such difficulty attends its application to the Atlantic islands.
It may fairly be asked, Why have Polynesian snail faunas failed to be enriched by the action of those means of dispersal which peopled the Atlantic islands? (1) Because in mid-Polynesia the distances between islands are great, and few snails are likely to survive the voyage, whatever the means of transport. Similarly, in the Atlantic we find no American snails in the Azores in spite of favoring currents. (2) There has no doubt been a certain amount of exchange between the island groups. It is only thus that we can explain the presence on several archipelagos of identical species. But this diffusion has been almost entirely restricted to small or minute species, such as Pupidæ, Realiidæ, etc. "With the exception of the Auriculidæ it [Partula hyalina] is the only species of Polynesian land shell of its size common to two or three distant groups of islands. As the above statement does not harmonize with the distribution of the various species of Polynesian shells as recorded by different authors, I will add that in Dr. Pfeiffer's last volume of his Mon. Heliceorum, where he enumerates 77 species of Polynesian (I exclude the Melanesian and Pelew) Partula, there exist 26 errors in localities. . . . All the species which are diffused over one or more groups are invariably minute shells."13
These observations by the most experienced of Polynesian naturalists agree with my results in studying the Bermuda fauna,14 and confirms the opinion that the whole Polynesian fauna cannot be due to drift or chance immigrants.
In the case of Bifidaria (Pupa pediculus), Opeas juneea, and probably some other widespread species, I think it extremely likely that they have been carried in vessels from island to island during the period of human occupancy; in some cases probably before the historic period. But there doubtless are a few cases where minute land snails have been carried from one archipelago to another by natural means, and over vast distances. We can hardly explain otherwise the scanty faunas of most of the "low" islands, to say [[p. 578]] nothing of the unquestionable occurrence of Tornatellina in the Galapagos,15 this genus being a stranger and alien in America.
In the southwestern Pacific, in and near the area where Forbes maps his "Antipodea" and Hedley his "Melanesian Plateau," comparatively recent groups have invaded a portion of what probably was part of the earlier Pacific land-mass. Some of the early fauna remains, such as Partula and Tornatellina, but merely as stragglers in a more powerful, more recent fauna, which seems to justify the hypothesis of an early or mid-tertiary land extending oceanward to the Fijis,16 such as Hedley and others advocate. Obviously later land connections have obscured the far earlier record in this area.
That the hypothetical Pacific continent was finally separated from any other land as early as the middle mesozoic would seem to be indicated by the absence of numerous families of land snails which had become fully differentiated by the end of the mesozoic; and it is evident that the portion north of the equator must have been isolated before the final separation of the remainder from the mainland, not only from the high differentiation of the Achatinellidæ, but because certain operculates and some groups of Zonitidæ, such as Trochomorpha, did not reach the Hawaiian area.
To the southward, it is likely that land extended from the Marquesas (whence granite and gneiss have been reported) and the Austral group to the Fijis or further, and northwestward possibly to the Caroline and Marianne groups, although it is obvious that these have been influenced by their proximity to the Oriental region, by drift or other causes, chiefly shown in the presence of Eulota, Diplommatina and Pupina, as well as certain lizards, etc.17 Where the Pacific continent may have touched Australo-Asiatic land is of course quite idle to inquire at present.18
[[p. 579]] It may be noted here that the distribution of ants and lizards, from which Baur argues, affords no support to the position I have taken, because the former, by having a winged stage, could be much more widely spread by wind than snails, and the distribution of recent Pacific lacertilia cannot possibly date to nearly so early an epoch as that of Polynesian land snails, but is rather traceable to later elevations, after the main mass of the ancient Pacific continent had been isolated or largely submerged.
Returning to the hypothesis of Hutton and Von Ihering, it remains to examine the evidence for a connection between the supposed mid or South Pacific continent and southern South America or "Archiplata." Prof. Hutton, in his temperate and judicious paper of 1896, says:
"The theory of a mesozoic South Pacific continent not only explains the origin of the Australian and South American marsupials, but also the almost simultaneous appearance of different Eutherian mammals in North and South America. We must suppose that this continent threw off first New Zealand, then Australia, then Chili, and finally disappeared under the waves. . . . [[p. 580]] At a later date, as I pointed out in my former papers, New Zealand must have formed part of a large island joined to New Caledonia, but not to Australia. This has lately been called Antipodea by Dr. Forbes, and the Melanesian Plateau by Mr. C. Hedley. Still later again, New Zealand must have stretched south and obtained its Antarctic fauna and flora from Patagonia through a number of islands."19
Notwithstanding the able arguments of Hutton and Von Ihering for a connection between Chili and a Pacific continent, it seems to me to involve grave, in fact insurmountable, difficulties. To have supported a marsupial fauna, to say nothing of Eutherian mammals, this continent must have persisted at least to near the end of the cretaceous. But if so, it is inconceivable that there should be no trace in Polynesia of characteristic land mollusks of South America, and none in South America of typical Polynesian groups. It is equally inconceivable that the Pacific continent completely disappeared under the waves about the end of the mesozoic, and that the present islands are of later appearance; for their faunas bear the stamp of a vastly greater antiquity, and (always excepting the "low" islands) have not the characteristics of "drift" faunas. The common elements of Australo-Zealandic and Archiplatan life, such as marsupials, Bulimulidæ, crawfishes, etc., are conspicuously absent from Polynesia, and may better, it seems to me, be accounted for by the much-discussed Antarctic route.20 I cannot trace any connection between the mid-Pacific continent and either of the Americas, nor with any continental mass whatever since the time of the advent of mammals.
One of the chief difficulties in the theory herein advanced is that many of the island groups, notably the Hawaiian, seem to be composed of only comparatively recent volcanic and coral rocks. In this connection Wichmann (as quoted by Baur) remarks: "No older massive rocks or sedimentary strata are known from the other "volcanic" groups of islands of the Pacific Ocean, and on some of these, for example, the Galapagos or the Sandwich Islands--it seems really to be made out that they have been built up by [[p. 581]] younger and recent volcanic masses. There is every possibility, and even probability, however, that older formations served as a fundament, the examination of which is prevented by the extensive covering."
Hedley urges the weighty objection that mesozoic plant forms should have survived on so ancient a land. Possibly the great facility with which seeds may be transported over sea allowed a new flora to displace the old one.
SUMMARY.
The hypothesis of a late palæozoic or early mesozoic mid-Pacific continent (upon the sunken heights of which the present island-masses, volcanic or coral, have been superposed) is advanced to account for the constitution of Polynesian land-snail faunas, which are shown to be (1) nearly homogeneous over vast areas, (2) composed of ancient types, with no admixture of the great series of modern families, and (3) not derivable from any tertiary or modern continental fauna or faunas in the sense Atlantic island faunas have been derived. The mollusca, land and marine, supply no evidence that this Pacific continent was ever connected with or faunally affected by the Americas, but emphatically deny such connection.
_________________________
Notes Appearing in the Original Work
1. Trans. New Zealand Institute,
XXIV, 1892, "On the Ancient Relations between New Zealand and South America."
[[on p. 568]]
2. Proc. Linn. Soc. of New
South Wales, 1896, p. 36, "Theoretical Explanations of the Distribution
of Southern Faunas." Professor Hutton has given so full a summary of the
principal literature bearing upon the subject, that I feel it unnecessary
to lengthen the present communication by a bibliography. Two strong papers
upon similar topics have appeared in the same periodical since Hutton's
paper--Deane's "President's Address," P. L. S. N. S. W. for 1896,
p. 821, and Hedley's "A Zoogeographic Scheme for the Mid-Pacific," 1899,
p. 391. [[on p. 568]]
3. American Naturalist,
XXXI, 1897. [[on p. 568]]
4. See these Proceedings,
p. 564, for a sketch of the classification of Pulmonates. The earlier
phylogeny of the main groups may be roughly represented by the following
diagram: [[on p. 570, directly preceding the diagram]]
5. In tabulating the genera I have
taken a few liberties with the Conchological Society's British list, in
subdividing their "Helix" group, and with Dr. von Moellendorff's Philippine
list in lumping some of his families of Holopoda. These changes
are made merely that the values of the groups used may be more uniform.
To count families and genera in the three lists as they stand would give
a false idea. [[on p. 571]]
6. Existing Ditremata may be looked
upon as excessively modified from a very old branch of Pulmonates. I cannot
accept Dr. von Ihering's theory of the primitive structure of Vaginulus,
or the homology of the Stylommatophore lung with a renal duct. [[on
p. 571, in the table]]
7. The Polynesian Zonitidæ
are all unspecialized genera, the Philippine mainly of more complex structure,
and the British and American partly so. [[on p.
571, in the table]]
8. This family is used here in
the conventional sense, synonymous with Fischer's Stenogyridæ.
We know little of the structure of many small members, but probably they
will group in two rather widely separated families. [[on
p. 571, in the table]]
9. These Proceedings,
p. 426. [[on p. 573]]
10. It seems characteristic of
islands that while specific differentiation usually proceeds apace, more
fundamental changes are retarded, or perhaps, more properly, are not induced;
and the faunæ lag behind those of the continents, old types lingering
on. [[on p. 573]]
11. In writing that "none of these
oceanic archipelagos present us with a single type which we may suppose
to have been preserved from mesozoic times" (l. c., p. 305),
Wallace makes a statement totally at variance with the nature of their
land mollusks. [[on p. 574]]
12. Pupina and Eulota
also occur in the Carolines. Flammulina will doubtless be recognized
in other island groups. [[on p. 576]]
13. A. J. Garrett, in Journ.
Acad. Nat. Sci. Phila., VIII, 396. [[on p.
577]]
14. Trans. Conn. Acad. Sci.,
X, 1900. [[on p. 577]]
15. See Dall, these Proceedings
for 1900, p. 95. Tornatellina chathamensis is the sole species
of Polynesian affinities known in the Galapagos. The supposed Endodonta
I would refer to Stobilops, and the Trochomorpha to
Guppya, both American groups. [[on p. 578]]
16. Cf. Baur, Amer.
Nat., 1897, p. 676, where the Batrachians and Reptiles of the Fijis
are discussed, with a summary of Wichmann's conclusions on their geology.
[[on p. 578]]
17. See v. Moellendorff, Land
Shells of the Caroline Islands, Journal of Malacology,
VII, pt. 5, 1900. Baur, Amer. Naturalist, 1897, p. 879, et
seq. [[on p. 578]]
18. The following extract from
a letter received from Mr. Charles Hedley contains suggestions of value
in this connection:
"In a recent paper on the
Caroline Island Tahitian shells by Moellendorff I have been struck by
the similarity to the Tahitian fauna. Can you make [[p.
579]] out any sequence between the Partula from North
and South Pacific? It would be interesting to learn that those of the
Carolines were a derivative of the Tahitian or the reverse.
"Now if you accept my theory
that the Marshall-Austral chain is not a phantom but a reality, an earth-fold
or line of weakness in the terrestrial crust, then we may suppose a former
more or less continuous land connection between Tahiti and the Carolines.
It would have served for the transmission of Garretia, Partula
and your other primitive snails. On its connection by drift with the Placostylus
land we are agreed. In the Gilbert and the Ellice Islands, as I have previously
said, the land sank, drowning the primitive fauna, rose again and was
repopulated by drift. I am inclined to think that the same fate overtook
Samoa. That, however, on rising pressed an earth-fold against the Fijian
massif and swelled into lofty mountains. That archipelago derived
Ostodes, etc., by drift from Fiji. Hence it lacked many characteristic
forms which survived in Tahiti.
"A grave objection
to your hypothesis, and one which deterred me from valuing Tahiti as continental
instead of oceanic, is that it proves too much. Although a palæozoic
continent is so far back as to be almost out of sight, still some other
forms ought to be found to support your snail fauna. I do not press the
fact that the insect fauna is extremely poor, since insects are held to
be of comparatively recent date. But I do press for collateral evidence
from the vegetation. I should like to see an old proteaceous genus, for
example, produced. Most of all I emphasize the absence of any peculiar
marine molluscs. I think that Clavella is peculiar, but I have
found none other. The argument that Nautilus, Onchidium
and the Polyplacophora are incapable of drift and though ancient forms
are conspicuously absent from Polynesia is worth careful attention. I
do not lean on the absence of fossiliferous beds, since circumstances
may be imagined under which they might have existed and disappeared."
[[on pp. 578-579]]
19. Proc. Linn. Soc. N. S. Wales,
1896, p. 46. [[on p. 580]]
20. The only Polynesian type in
America known to me is Tornatellina, in the Galapagos, as already
noted. A minute straggler or two may reasonably be expected. The present
fauna of "Archiplata" is as remote as any on earth from that of Polynesia.
[[on p. 580]]
*
*
*
*
*