
http://people.wku.edu/charles.smith/biogeog/HARR1924.htm
The Migration Route of the Australian Marsupial
Fauna. Presidential Address.
by Launcelot Harrison (1924)
Editor Charles H. Smith's Note: Original pagination
indicated within double brackets. Notes are numbered sequentially and
grouped at the end, with the page(s) they originally appeared at the bottom
of given within double brackets. The paper is presented complete, the
presence of several sequences of " . . . ." notwithstanding. Citation:
Australian Zoologist 3 (1924): 247-263.
[[p. 247]] It may
justifiably be said that a majority of Australian zoologists, as well
as many botanists, believe that Australia has in past time been connected
by land with South America, and that a considerable proportion, both of
its plants and animals, has been derived from the latter Continent by
land migration. There is a good deal of evidence for the extension of
land masses well into the eastern Pacific, which joined up with New Zealand
to the south, and with Papua to the north. But there is a vast stretch
of deep ocean between the easternmost of those islands and submarine banks,
which afford indication of former land extensions, and the coast of South
America, and a land bridge in this direction would involve a radical change
of level in the floor of the Pacific. Such a change is not acceptable
to geologists, though that need not place it outside the bounds of possibility.
But Australian opinion favours a connection through former extensions
of the Antarctic Continent, linking up with South America, Australia,
and Africa, and there is a widespread belief that much of our fauna was
received from America along this route.
In recent years, however, several important attacks have been launched against this hypothesis. The first of these which I propose to consider is that of W. D. Matthew (1915), who, in an extremely able and comprehensive survey of the distribution of mammals from the Tertiary onwards, argues for a Holarctic dispersal centre for all the major mammalian groups. Matthew correlates mammalian migration with the migration of climates, adopting the view that climatic change has been of the nature of alternating periods of glaciation followed by aridity and epi-continental flooding accompanied by moist climates and the development of rain forest. This view is that of the American school of geologists, led by Chamberlin. With the broad features of Matthew's thesis one can quite agree, but when he endeavours to lay down one way, and one way only, for mammalian distribution, be becomes involved in difficulties which require measures even more drastic than inter-continental bridges for their solution. Matthew claims a northern derivation for the Australian marsupials, and incidentally considers the distribution of crayfish and frogs, two groups which have been used extensively for the purpose of arguing a southern connection between Australia and South America.
More recently G. K. Noble (1922) and E. R. Dunn (1923) have discussed the distribution of frogs, both claiming a northern origin for the Australian Hylidae and Leptodactylidae. As the distribution of these families in Australia and South America has hitherto been one of the strongest arguments in favour of a former land connection, it is necessary that careful consideration should be given to their views. Maynard M. Metcalf (1923), on the other hand, brings forward very strong additional evidence, of a circumstantial nature, no doubt, but in accordance with the writer's views on the distribution of parasites, of the direct relationship between Australian and South American forms.
[[p. 248]] Finally H. A. Longman (1924), has advocated a northern origin for the marsupials, basing his belief upon the variety of forms found in the islands to the north of Australia, and upon the past distribution of marsupials in Queensland.
Matthew's views are perhaps of most importance, and it is of interest to examine the general nature of the theses upon which they rest. These must be presented in very summary form, and it is difficult to be entirely fair under such circumstances, but I have at least made an honest endeavour to present them impartially. Matthew states that climatic change is the chief known cause of the distribution of land vertebrates, and that the principal lines of migration were radial from Holarctic centres. The geographic changes required to explain distribution are not extensive, and, for the most part, do not affect the permanence of ocean basins. The idea of alternations of moist and uniform with arid and zonal climates accords with the facts of distribution. Land bridges over the present deep ocean are improbable and unnecessary.
He adopts Chamberlin's views on the alternation of climates between extremes of warm moist tropical climates, resulting from prolonged base-level erosion and overflow of large continental areas by shallow seas; and cold arid zonal climates resulting from re-establishment of isostatic balance, with the expansion of continental areas to the limits of their continental shelves. As regards their geological residues, the former are marked by limestone and coal, the latter by thick barren formations, culminating in a great extension of glaciers from boreal and high mountain areas. He limits the possible emergence of continental land masses to their existing continental shelves, except along those lines of weakness or instability which have been subject to folding, with great changes of level. He indicates the essential distinction between the faunae of these alternating climatic phases. The former phase implies isolated continents, with provincial land fauna, which, owing to abundant food supply and easy conditions of life, would tend to be sluggish in the matter of evolution; while the latter implies, under conditions of cold and aridity, a stimulus to migration and differentiation along the now connected land masses, with cosmopolitan fauna, the more primitive forms being pushed southwards.
As regards the time of dispersal, Matthew writes (l.c., p. 179):--"With a clearer perspective of geologic time and far more exact records, it is clear that most of this deployment and dispersal of the mammalian races has taken place since the Eocene epoch of the Tertiary, although remnants of an older dispersal on the same lines are probably traceable in the present habitat of monotremes, marsupials, and primitive insectivores."
Matthew's interpretation of supposed exceptions to his general rule is not so satisfactory. He writes (l.c., p. 179):--"There has been a disposition in recent years among students of geographical distribution to lay weight upon certain apparent exceptions1 to this general rule, where the geological record has not yet afforded evidence to support the northerly origin of certain groups2 now limited to the southern continents or to the tropics and to infer various equatorial or southern continental connections during or previous to the Tertiary, in order to account for these exceptions. To these hypotheses, there are several objections:--
"(1) The evidence for the general permanence of the great ocean basins and their maintenance formerly, as now, by isostatic balance is very strong and direct, and before allowing any exceptions, we should be very sure that no other explanation will serve.
"(2) The instances adduced in favor of former equatorial or southern connections are distinctly exceptional cases in the faunae, which may, in all the cases [[p. 249]] I have examined, be accounted for by appealing to the imperfection of the geologic record, by parallelism or by the rare accidents of oversea transportation.
"(3) The existence of such land bridges would present the opportunity for migration of other parts or of the whole of certain faunae, which has evidently not occurred. I can see no good reason why the only animals which availed themselves of such continental bridges should be the ones which might be accounted for in other ways, while those which would furnish conclusive proof are invariably absent.
"(4) Many students of geographic distribution proceed on what appear to me to be wholly false premises. They assume that the habitat of the most primitive living member of a race is the original habitat of the race, the most advanced forms inhabiting the limit of its migration. It seems to me that we should assume directly the reverse of this."
The words I have italicised above show how Matthew's argument descends on occasion to dialectic. The facts are that in our present state of knowledge the distribution of certain groups does not conform with Matthew's views. By calling these "apparent exceptions" and introducing the word "yet" into the next clause, Matthew implies that further palaeontological discovery will bring these groups into line with his ideas, an assumption for which there is, at present, no justification whatever.
As regards paragraph (1), we may admit that there is very strong evidence for the permanent nature of the great ocean basins, but many of the land bridges which have from time to time been suggested would not interfere to any extent with that permanency. A glance at the south polar projection shown in Matthew's, Fig. 2 (p. 176) will show that no great degree of elevation of the Antarctic hemisphere would bring Australia, New Zealand, South America, and Africa into connection with an extended Antarctica.
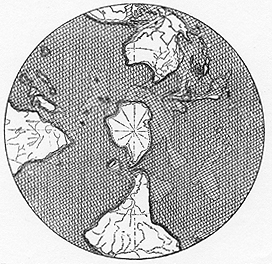
Fig. 1.--The Southern Hemisphere on a south polar projection. After Matthew.
[[p. 250]] Matthew's argument (p. 189) that--"The permanency of the continental platforms is indicated by the absence of abyssal deposits in their sedimentary succession wherever this has been adequately studied," only holds in so far that geologists are somewhat reluctant to apply their principle of uniformity to such deposits when they are observed. He mentions chalk found in shallow water, which may be possible, but even radiolarian oozes, hardened into cherts, are claimed as shallow water deposits, a claim which is entirely untenable. The radiolarian cherts of north-eastern New South Wales indicate a deposition at 2,000 fathoms or more just as surely as do the radiolarian oozes to-day. Yet these are stated to be of shallow water origin on the ground that similar cherts have had the same claim made for them in other parts of the world. A radiolarian deposit, free from calcareous remains, cannot be formed except in ocean abysses; and such a deposit found on an existing continental platform indicates a very great change of level in past time. There is evidence for a relative permanence of continental platforms and ocean basins, but the claim for absolute permanence cannot be maintained.
Paragraphs (2) and (3) I find difficult to understand. Some of the statements may apply to a fraction of the land bridges which have from time to time been hypothecated, or, possibly, even to a majority of them, but I am here concerned only with the past land connections of Australia, and to these I cannot see that they apply. A great deal of the Australian flora, almost the whole of the mammalian and amphibian faunae, and a long list of genera, families, and even orders of other faunal groups find their nearest relatives in South America, and it cannot be said that these are "exceptional cases in the faunae." The three ways by which Matthew would account for this condition are all of them ways for which no positive and direct evidence can exist. The objection that--"The existence of such land bridges would present the opportunity for migration of other parts or of the whole of certain faunae, which has evidently not occurred" falls entirely, for it is just this that a majority of Australian zoologists holds to have occurred. It is, of course, obvious that only co-existent groups could make use of any given land bridge, unless it persisted through a very long period, a consideration that would appear to have evaded Matthew, or been evaded by him, in some of his detailed arguments.
The last sentence of paragraph (3) has no meaning for me. When a certain proportion of the members of every animal Phylum found in Australia shows South American affinities, which, however, I do not concede that Matthew has accounted for in other ways, it would be interesting to learn just what other groups of South American animals, if discovered in Australia by some happy chance, would afford evidence more convincing than that already provided, or less easily "accounted for in other ways."
With the broader features of Matthew's principles of dispersal one may agree, but he himself has had to admit exceptions, and the single Holarctic dispersal centre which apparently holds good for most mammalian migrations during the Tertiary will not hold for all. The outstanding feature of his masterly analysis of these migrations from the viewpoint of the present enquiry lies in the emphatic re-statement of the isolation of Australia from the northern land masses. We know quite well that the placental mammals failed to reach Australia, but the fact seems to take on a greater significance as we find it restated in the analysis of each successive group. This isolation Matthew admits (p. 232) to have extended through the whole of the Tertiary.
Matthew's views on the dispersal of marsupials (pp. 262-269) may be summarised as follows:--The present distribution is almost limited to the Australian and Austromalayan region. A single unspecialised group, the opossums, close to
[[p. 251]]
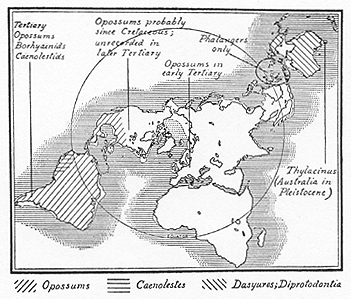
Fig. 2.--The distribution of marsupials. After Matthew.
the primitive type from which all marsupials have developed, survives in the Neotropical and ranges northwards into the Sonoran sub-region of North America. Caenolestes is not a diprotodont, but an American radiation from the polyprotodonta. The first remains which seem definitely those of marsupials occur in the uppermost Cretaceous of Wyoming. No marsupials have been recognised in the Holarctic basal Eocene. Polyprotodonts definitely occur in the Lower and Middle Eocene of Wyoming, in the Oligocene of Colorado and in the Upper Eocene to lower Miocene of France and Germany. In the southern continents, they assumed a much more important position. The adaptive radiation in South America was not so marked as in Australia, owing to the fact that the former was invaded by certain primitive placental groups before it was cut off from North America. The South American Borhyaenids show homoplastic resemblance to, and not relationship with, Thylacinus, so that this evidence for Antarctic connection fails. Caenolestes and the Epanorthids are modified polyprotodonts, not, as was at first supposed, primitive diprotodonts.
P. 267--"In view of the great amount of adaptive divergence seen in the various Pleistocene and modern genera of Australian Diprotodonta, the origin of the suborder in Australasia or its earliest invasion of that zoological region, must be dated far back in the Tertiary. On our present evidence it may well be regarded as wholly autochthonic, derived from early Tertiary or possibly from late Mesozoic polyprotodonts. Nevertheless, in view of the defectiveness of the Mesozoic record, where we should chiefly expect to find this group, if anywhere in the North, and the presumable rarity of Tertiary survivors, there is nothing unlikely in the view that they originated primarily in the North like their polyprotodont and allotherian relatives and were driven southward with the former group and somewhat more thoroughly extinguished in the north, while in [[p. 252]] Australia they blossomed out into a great adaptive expansion paralleling the absent ungulate mammals."3
There is no clear record of Didelphids in the Miocene or Pliocene of North America, none after the Lower Miocene in Western Europe, nor in the later Tertiaries of India or China.
Such is the body of fact upon which Matthew bases the opinions I have italicised, and that opinion goes no further than to claim that there is nothing unlikely in the view of northern derivation. It seems to me, however, that this conclusion is open to criticism on sound grounds. Admitting the imperfection of the geological record, the actual evidence derivable from it is that the earliest Holarctic marsupials occur in the late Cretaceous; that none are known from the Palaearctic after the Lower Miocene; and that none at all are known from south-eastern Asia. It is possible to argue upon this basis that North America formed the centre of dispersal, though, despite disagreements about the correlation of the South American horizons, it seems certain enough that there was a well differentiated marsupial fauna in that continent at the beginning of the Tertiary, and more probable that this evolved in situ from Mesozoic ancestors. At any rate, more marsupials, fossil and recent, are known from South America than from any other part of the world outside Australia, and there is little evidence for any considerable development of even the most primitive marsupials in the Palaearctic land mass. Any argument for the incompleteness of the geological record in Asia applies with equal cogency to Antarctica. What we do know about the latter continent palaeontologically may be quoted from David and Priestley (1914, pp. 313-314), who, after pointing out that the five plant genera Sagenopteris, Thinnfeldia, Cladophlebis, Pterophyllum, Otozamites are common to Antarctica, and Indo-Australia, while the second and third occur in South Africa, and the second and fourth in the Argentine, write:--
"The close affinity of the flora to that of Australia and of India, shown on this list, implies a land connection between Antarctica and Australia in Jurassic or Trias-Jura time.
"At Snow Hill Island abundance of Cretaceous fossils, including numerous Ammonites, were collected by Dr. Nordenskjöld's Expedition. These fossils imply the existence of a mild climate, with comparatively warm ocean currents in the neighbourhood of Snow Hill Island in Cretaceous time. The fossil plants Araucaria, Fagus, etc., near to A. braziliensis, unearthed by the Nordenskjöld Expedition at Seymour Island, adjoining Snow Hill Island on the north-east, prove that these mild weather conditions were further prolonged into some part of Tertiary time, probably Oligocene or Lower Miocene. In marine strata, also of Tertiary age, and considered by Wilckens to belong to Upper Oligocene or Lower Miocene, this expedition found numerous bird bones, since referred to five new genera of penguins, besides two vertebrae of a big mammal referred to the genus Zeuglodon. It is very interesting to note that remains of a large penguin have been obtained in New Zealand in the Oamaru beds, which are considered to be of Eocene age."
Matthew dismisses Gondwana Land as being outside the scope of his discussion, but to leave this question of Antarctic connections without discussing the Mesozoic floral distribution is to offer Hamlet without the prince. In the first place, we have evidence that Antarctica possessed a mild climate and a rain forest flora throughout the Mesozoic and into the Tertiary as far, perhaps, as Lower Miocene. Secondly, there is demonstrated the existence of at least two distinct florae, a Mesozoic Gondwana flora, and a Tertiary flora with South American affinities, including Araucarias and small-leaved beeches. This latter [[p. 253]] flora occurs in both New Zealand and Australia. Cheeseman (1909) has discussed the floral relationships of Fuegia with the ring of circum-polar sub-antarctic islands and with New Zealand, Australia, and South Africa. While he is dubious about actual land connections, he considers that these probably were extensions of the existing land masses, which brought them in closer proximity to one another, and he can explain the existence of a proportion of the sub-antarctic flora only on the ground that it has been derived from the Antarctic continent.
If Antarctica possessed a rain forest flora through the length of time indicated, from the Jurassic into the Tertiary, it seems equally certain that it must have possessed a fauna. Some happy chance may in the future throw some palaeontological light upon the nature of this fauna, but it seems most likely that the mantle of ice and snow which covers the continent will keep the matter a mystery for us. But when we know quite definitely that certain kinds of plants occurred there, which are still found in South America, in New Zealand, and in Australia, and when we see that many kinds of animals are common to these three regions, and afford no evidence whatever of northern origin, despite the fact that we have not found them fossil in Antarctica, it seems to me that we are well justified in assuming their existence there in times past. I have not time to traverse the large body of evidence relating to the invertebrates, but in almost every group there are members which can most plausibly be accounted for by a southern origin. Benham (1909) has put forward a particularly good case for the Oligochaetes, which also holds to some extent for groups like the land planarians and land nemerteans, all creatures for which it is difficult to imagine a drift transportation. Australian systematic zoology teems with forms exhibiting South American affinities. Curiously enough, many of these forms find their nearest relatives in Chili, but this is probably due to the similar nature of the physical environment, and does not necessarily argue for a direct trans-Pacific connection. Men who have been most intimately acquainted with the invertebrate fauna, Hutton and Benham in New Zealand, Haswell and Hedley in Australia, have all agreed upon the necessity of Antarctic connections.
It is obvious that these could not have been contemporaneous. Marsupials did not reach New Zealand, and it is still a somewhat doubtful question whether any reached Africa. Benham's evidence (loc. cit.) from earthworms would indicate that connection with New Zealand post-dated that with Australia, and may have occurred after our supposed Antarctic marsupials had been killed off by increasing cold. But the data available at present are not sufficient to allow of putting a period to any of these hypothetical land connections. All we can say is that Africa and Australia, common possessors of a Proteacean flora, would seem at some time to have been connected, probably through the south; that there is some evidence for a separate connection with Madagascar; and also that Australia, New Zealand, and South America would appear to have had connections with Antarctica, though not contemporaneously. Matthew would consider the related faunae and florae upon which these hypotheses are based as residuals of a once cosmopolitan distribution, driven southward along the three great peninsulas. I would reply that there is no actual evidence for this view, and that the weight of probability is against it. Apart from the mere matter of probability, there is evidence of a kind which may not be generally acceptable, but which I believe to be absolutely reliable, to the discussion of which we shall presently come.
One argument for actual land connections as against the mere narrowing of intervening seas, may be drawn from Australia itself. Australia has been isolated from the north since the beginning of the Tertiary, or, possibly, since late Mesozoic time. To the north lies a group of islands which have suffered geographical [[p. 254]] vicissitudes not yet fully worked out. The lines of Wallace and Weber may not be absolute faunistic boundaries, but it seems likely that they have a significance, and that Wallace's line does, in a general way, mark the line of separation of Austro-Malaya from Asia, a separation that most probably took place in the Mesozoic. The only indication that any of the most primitive mammals reached Australia lies in the fact that the existing monotremes are still found there, and these are certainly derivatives of a very primitive mammalian stock. Unfortunately we know nothing about the history of the monotremes. Various fossil remains, particularly in South America, have been attributed to the group, but none has been proved definitely to belong to it. The genus Ornithorhynchus has existed unchanged in Australia since the Pliocene, and its specialisation from a primitive stock must have been a long business. It seems probable that the ancestors of the existing monotremes entered Australia from the north during the Mesozoic, possibly in the Jurassic, and that soon after their arrival the land way was cut. Had this not been the case, other primitive mammalian stocks would surely have entered. No evidence for such invasion has been found, but little weight can be attached to this, for there is equally no evidence for the invasion of the ancestral monotremes, which must have occurred.
In any case, it seems clear that from the beginning of the Tertiary no mammals reached Australia from the north. The point I want to make is that, although Australia is almost connected by a chain of islands, with Malaysia, there has been comparatively little accidental transportation of fauna. The stocking of the East Indian islands appears, according to the latest work of the Dutch geologists, to have been accomplished through alternate joining up and severance of land masses. In this sub-region, which offers conditions peculiarly favourable to accidental transportation, there is very little evidence for its occurrence, except that afforded by a few rodents, other than which no terrestrial placental has been able to reach Australia. Now if porcupines can be rafted from South America to Africa as Matthew suggests (certainly with some diffidence, but without any suggestion as to how the raft was provisioned for the voyage) and if this sort of thing happens generally, even though rarely, why has it not happened to Australia? Since it has not happened here, although conditions are most favourable, we are surely justified in concluding that it does not happen elsewhere, and that land mammals require land bridges for their migrations.
The question remains--Was the land bridge by which the marsupials reached Australia to the north, or to the south? In America, marsupials reach to about the fortieth parallel of north latitude. In Austro-Malaya, no marsupial reaches the equator. Accepting Matthew's view that the northern phalangers are a radiation from a Papuan centre of dispersal, they have been prevented by a hitherto impassable barrier from passing to the north and west of Celebes. But the polyprotodonts have an even more restricted range, extending only as far as Ceram. This is not what one would expect if their primitive ancestors came by way of the Moluccas. It is a simple matter to assume that these have become extinct throughout south-eastern Asia and the greater number of the East Indian islands. But why should they have become extinct? The physical environment is in every way suitable, and small cryptozoic marsupials are not in the least likely to have been killed out. It seems much more probable that they were never there, a probability that is much strengthened by the similarity of distribution in frogs of the families Hylidae and Leptodactylidae which are discussed below. Accepting all the facts that Matthew has so carefully put together, I can still see no reason for the deductions he makes from them. It is true that certain pieces of evidence that were formerly used to support the Antarctic connection have to be resigned. Caenolestes is not a primitive diprotodont, Borhyaena is not a Thylacinid, Galaxias breeds in the sea. But these things do not appreciably [[p. 255]] lessen the evidence derived from so many groups of affinities most easily explained by Antarctic connections.
Longman (1924) analyses the present distribution of Australian marsupials and points to the considerable number of species of Peramelidae and Dasyuridae, known now to occur in North Queensland and Papua, as evidence of northern derivation. According to Matthew's views, however, which hold good so generally for Holarctic dispersal centres, it is just precisely this kind of distribution that we should expect if these admittedly primitive marsupials had become dispersed from a southern centre. In fact, Matthew, admitting the dispersal of the Diprotodonts from an Australian centre, writes (1915, p. 263):--"The Phalangers of the Austromalayan islands are regarded as marginal types from an Australian dispersal centre." Of the Polyprotodonts, however, Matthew (l.c., p. 295) states that they have not--"entirely disappeared from the East Indian islands." Since the Phalangers and the more primitive Polyprotodonts have, in general, the same total marginal distribution, it is surely not reasonable to interpret this distribution as evidence of Australian dispersal in one instance, and of northern dispersal in the other. The primitive Polyprotodonts are generally assumed to have been derived from primitive Didelphids, and may have had either a Holarctic or a Neotropical derivation. But there is no evidence at present derivable from marsupials themselves, whether fossil or recent, as to the actual source of origin.
The nature of the Papuan sub-region introduces a further factor which has not been considered by Longman, but which must have been of great importance in stimulating the formation of species. Throughout this sub-region we have high mountains and dense rain-forests, and a considerable amount of geological change. That a number of arboricolons and small terrestrial species should have become differentiated is not at all remarkable. The evidence from other sources makes it clear that Papua has served as a dispersal centre for a number of local groups, and it is quite probable that a limited dispersal of Phalangers may have taken place in this way. Nevertheless it seems equally probable that there was an earlier dispersal from an Australian centre.
Much of Longman's argument is directed against the existence of the diprotodont condition in South American marsupials. But before the supposed American diprotodonts were discovered it was considered that this condition originated in Australia. We have lost nothing through the discovery of the correct affinities of these American forms. Syndactyly is an adaptation to arboreal life, and may as Lönnberg (1921, p. 96) suggests, have occurred independently in Australian polyprotodonts and diprotodonts.
Longman writes (loc. cit., p. 12):--"The assumption of a continent in the Antarctic in early Tertiary times"--as if this itself were hypothetical. But the continent was there from the Palaeozoic, had a Mesozoic rain-forest flora, and a Tertiary flora also. Moreover, it is still there to-day, although its flora has been reduced to a single phanerogam and a few cryptogams, owing to intensive glaciation.
Longman's conclusions are:--
1. The "fundamental consideration" that marsupials were once widely distributed in Holarctic regions. This, of itself, proves nothing.
2. The diversity of marsupial genera in North Australia, and Papua. This is only natural, but has no bearing on the question of origin.
3. The evolution of the Australian marsupials has taken place in Australia. I am not aware that this has ever been disputed.
4. Wynyardia is not akin to the Caenolestids. This is immaterial. What is of importance is that Wynyardia seems to be in some respects intermediate between the polyprotodonts and the diprotodonts.
5. The diprotodonts have nothing in common with South American forms. [[p. 256]] Since they are presumed to have been derived from polyprotodonts in Australia, there is no reason why they should.
6. All living families of marsupials were well differentiated early in the Tertiary. This only makes a Cretaceous migration to Australia the more probable.
7. The rich development of Peramelidae in the Austro-Malayan region. This development is almost entirely Papuan, and it is well-established that Papua has been joined to Australia.
8. The primitive nature of syndactylism. Whether the view suggested above be correct or not, syndactylism has arisen in Australia, and does not affect the general question of origin.
Matthew (l.c., pp. 294-296) discusses the dispersal of Amphibia, and the following quotations are pertinent to our discussion:--
"Among the modern families the Cystignathidae4 are chiefly Australasian and Neotropical, but a few are still5 found in North America. This distribution parallels that of the polyprotodont marsupials, except that the latter have not reached New Zealand or the Antilles, or entirely disappeared from6 the East Indian islands.
"The genus Bufo has failed to reach Australasia, Madagascar or New Zealand, but is replaced in Australia by a (more primitive?) member of the family. The Hylidae are to-day chiefly South American and Australian, but a few members still7 inhabit North America. They are not found in Africa or the Oriental region, where it seems reasonable to suppose that they have been displaced by the true frogs (Ranidae), peculiarly varied and abundant in these regions. The Ranidae, like the Bufonidae, represent a less ancient dispersal, probably from a southern Palaearctic or Oriental centre, since they have reached northern Australia on one side and north-western South America on the other, and, while they have reached Madagascar and the Solomon Islands, they have failed to reach the Antilles."
The words italicised exhibit again the partisan twist which Matthew gives to much of his argument. In each case there is an inference or implication added to the statement of fact. There is no evidence whatever that any greater number of Hylids or Leptodactylids ever inhabited North America, and the implication that they are residuals of a larger fauna when it seems much more probable that they are fairly recently derived from the south, and have never greatly extended further northwards and westwards, is unjustifiable. The same holds for the statement that the polyprotodont marsupials have not entirely disappeared from the East Indian islands. There is not one jot of positive evidence to indicate that any polyprotodont marsupial has ever disappeared from any East Indian island. Had they ever existed in the islands not at present tenanted by them, all the probabilities are that they would have persisted to the present, for these islands afford an environment similar to that provided by neighbouring islands in which they contrive to flourish and even undergo adaptive radiation.
Noble (1922) has adopted and expanded Matthew's views. He writes (l.c., p. 63):--
"If the recent herpetological literature is critically examined, it will appear that to-day there is no need for the antarctic continents or mid-Atlantic land bridges which have been revived from time to time to account for the distribution of the Salientia. Perhaps the most recent of these revivals is that of Metcalf (1921a) who has brought some additional evidence derived from a study [[p. 257]] of the Opalinid parasites of the Salientia. There seems to me to be little need for the land bridges Metcalf has revived. Hylids and 'leptodactylids' occur in the Ethiopian region. We have merged the 'leptodactylids' into the Bufonidae, while the hylids have almost certainly evolved from this bufonid stock." Noble assumes a polar origin for the Hylidae, and Leptodactylidae, on the ground that the Brevicipitidae, Ranidae, and Bufonidae, 'have almost certainly gained access to Africa from the north.' He writes further (p. 69):--
"The genus Hyla possesses nearly a world-wide distribution, except for a great gap in the Indo-Oriental region. Formerly considerable emphasis was placed on this gap in its present distribution. This, together with the abundance of the genus in the South American and Australian regions, seemed to lend some support to the Gondwana land hypothesis. But to-day we know of one true Hyla from the Ethiopian region and three toothed bufonids. The latter are all referable to the single genus Heleophryne. It thus seems most likely that the genus Hyla originated in the north and pushed southward into Africa and the Oriental region. Finally, when the genus gained access to Australia, it died out in most of the more northern regions except in Papua and some of the East Indian islands where it is still abundant."
Noble has united the Bufonidae and Leptodactylidae, calling them all Bufonids. This action has not had time to draw upon itself the requisite criticism which will decide whether it be well founded or no. It does, however, enable Noble to claim a continuity of Bufonidae from Asia into Australia which, on closer analysis, does not exist. Typical northern Bufonids of the genus Bufo have not been able to penetrate to the east or south of Celebes. The Australian Leptodactylids, the essential characters of which are not in any way altered by calling them Bufonids, have not succeeded in penetrating to the north or west of Papua, in which island they are represented by one or two species only; and their closest affinities would still seem to lie with South American forms. The occurrence of Heleophryne in Africa would not be inconsistent with Antarctic derivation, probably after Australia was cut off from its Antarctic connection. There is evidence for this African connection shown by the Proteaceae among plants. Moreover, Heleophryne and the reported Pseudophryne of Africa may be wrongly determined as Bufonids. I am not, however, concerned about Africa at present, and prefer to consider the Hylidae only, since they do not raise any question of African relationships. Before proceeding to this, however, a few more passages from Noble's paper must be added, though they merely reiterate the same arguments.
He writes (p. 70):--"The fact that numerous toothed bufonids occur in Australia and in South America has for many years lent some support to the Antarctic continent theory. We have already stated above that this recent discovery of toothed bufonids in Africa together with a recognition of the close affinity of toothed and toothless forms removes much of the argument in favor of this view. For years the distribution of hylids has been used as supplementary evidence in favor of a former South American-Australian land connection. . . . The distribution of the genus Hyla is very difficult to explain. We have already remarked on the discovery of a Hyla in Abyssinia. The genus is now known from all the major zoological regions except for a remarkable hiatus, extending from the Philippines, Borneo, Celebes, Siam, and the Malay Archipelago through India. Several of the Asiatic Hylas encroach upon the border of this region (as in Burma), but these are readily recognisable as ectogenetic forms. Why this Indo-Oriental region should be a closed territory to Hyla is not known, but I do not believe there is good reason to assume that it has always been a closed territory, that the genus has never existed in any of this region. If we overlook for a moment the great gap in an almost world-wide distribution for the genus Hyla [[p. 258]] . . . . it is much simpler to account for the present distributions . . . . by assuming a northern origin and later migration southward by three routes, the first leading into South America, the second into Africa, and the third into the East Indies and Australia, than to assume land connections between South America and Australia when we would have still to account for the Hylas in Africa and northern Asia."
I would suggest that the reader of Noble's argument who was totally unacquainted with Anuran distribution would get a hopelessly wrong idea of it from the selection of facts which he sets out. The alleged world-wide distribution of Hylidae is perhaps verbally true, but the distribution in the Oriental, Palaearctic, and Ethiopian regions, much more than half the world, is limited to two species, one of which is divided into half a dozen varieties, while the other appears to be only doubtfully known. These two species, according to Metcalf and Noble, are closely related to American forms. In the case of Hyla arborea and its races it seems certain that it migrated from America via Alaska. About H. wachei so little is known that it is difficult to base any conclusion upon it. If Noble's suggestion (l.c., p. 65) be well founded, it is most closely related to the maxima group of northern South America, and may have followed the same route as H. arborea.
Dunn (1923) does little more than reiterate Noble's conclusions. Noble quotes Metcalf (1923), but appears to allow little weight to the evidence the latter adduces from the distribution of Opalinid parasites of frogs. Metcalf finds that Hyla arborea and its subspecies harbour Opalinids belonging to a modern group, which he believes to have originated in North America. The Australian Hylas are tenanted by the primitive genus Protoopalina, while the "narrow Opalinas" found in American and Holarctic Hylas are thought by Metcalf to have been brought about through the reactions of these hosts upon the "broad Opalinas" of Rana and Bufo.
Metcalf concludes (1923, p. 394):--"No multinucleated Opalinids are known from Australasia. If the Australian Hylas had entered from Asia they should have brought multinucleated Cepedeas or Opalinas, such as are found in Nearctic Hylids, unless they left Asia before the evolution and presence there of these multinucleated genera. All the indications seem distinctly against the hypothesis of the Australasian Hylids having entered Australasia from the north."
The evidence is not conclusive, as Metcalf admits, but it is strongly supported by that derived from the parasites of the Leptodactylids. The Opalinid genus Zelleriella is found only in Australia and southern America. As Metcalf points out, if the Australian and South American Leptodactylids were of northern origin, then, on the basis of what happens with other Opalinids, Zelleriella would still occur in some Anuran hosts in Holarctica, which it does not. The distribution of Opalinid parasites seems to me conclusive for southern derivation of the host groups.
There may of course be objection taken to this host-parasite method of arriving at conclusions of such far reaching importance. But I believe it to be well justified. I first proposed in 1911 that the Mallophagan parasites of birds might settle some vexed questions in bird phylogeny, and have used them to this end in several subsequent papers. In 1915, I put forward the general thesis "that in the case of total obligate parasites, closely related parasites will be found to occur upon phyletically connected hosts, without regard to other ecologic conditions. As the state of evolution of the parasite will be less advanced than that of the host, it follows as a corollary: That a study of such parasites may give valuable indications as to host phylogeny." My predecessor in the chair I now occupy, S. J. Johnston, tested my proposition for Trematoda and Cestoda (Johnston, 1913, 1914) and found that it held good. Metcalf quite independently found the [[p. 259]] value of the method with Opalinids. I shall make a further new application of it before I conclude this address.
Since it is with the marsupial fauna that we are mainly concerned, what of the parasites of the marsupials themselves? S. J. Johnston (1913, p. 278), writes:--"The trematode parasites from Australian marsupials are very interesting in this connexion. Two species of Harmostomum from Dasyurus and Perameles are very closely related to H. opisthotrias Lutz from an American Didelphys, so closely related that I am convinced that they must be considered as being derived from common ancestors. They thus afford some pretty convincing circumstantial evidence of the phylogenetic relationship of the Australian and South American marsupials.
"No less interesting are two new species of flukes, one from Dasyurus and the other from the platypus, which have been described in Part 4 of the Proceedings of the Linn. Soc. N.S.W. for 1912, as representatives of a subfamily intermediate in position between the Fasciolinae, flukes typically parasitic in the higher mammals and the Psilostominae parasitic in reptiles and birds."
Certain evidence from Cestodes (S. J. Johnston, 1914, p. 4) points also to the genetic affinity of Australian and American marsupials, but neither they nor the Trematodes help us in the matter of migration routes. The Mallophagan ectoparasites, however, may afford assistance. I wrote (1922, p. 154) apropos of a Mallophagan parasite of an American marsupial, Peramys:--"Mallophaga from Australian marsupials are contained in a family, the Boopidae, which finds its closest relations in the Gyropidae, a family found upon certain South American rodents. Certain South American rodents also harbour the two contained species of a third family, the Trimenoponidae. With the exception of these three small groups, all mammalian Mallophaga belong to the widely different family Trichodectidae, which is placed in a distinct super-family.
"Believing as I do that Mallophagan parasites afford valuable indications as to the genetic relationships of their hosts, I have always been puzzled by this distribution. That the marsupials of Australia should not carry the same kinds of parasites as the Eutherian mammals is reasonable enough. But, apart from marsupials, I should have expected all other mammalian Mallophaga to belong to the Trichodectidae. Hence the occurrence of two small, but distinct, families, not upon rodents in general, nor even upon American rodents in general, but on a limited number of South American rodent species, families which showed, moreover, some relationship with the Boopidae, but differed from all other Mallophaga, was difficult to reconcile with my ideas.
"The explanation would appear to be that such Ambyceran Mallophaga as occur on South American rodents have been migrants in the past from the marsupial stock. The new genus which I describe from a South American marsupial must be placed in the Trimenoponidae, but shows some marked features of resemblance to the Boopidae, and some points of contact with the Gyropidae. It is, of course, no use trying to base definite conclusions on a single marsupial-infesting species, but it seems likely that, when more information is available concerning the Mallophagan parasites of American rodents and marsupials, the suggestion thrown out here may be upheld. It is also possible that the discovery of further connecting forms will make it advisable to unite these three anomalous groups under one family name."
Curiously enough, further information was soon forthcoming, for my paper was anticipated by just a fortnight by G. F. Ferris (1922) whose communication not only bore the same title as my own, but described my species, so that my Acanthomenopon horridum falls to Ferris' Cummingsia peramydis. Ferris described also a genus Harrisonia from an Octodont rodent, and a second species [[p. 260]] of Cummingsia from Caenolestes, and the new forms which he has brought to light show obvious affinities with the Boopidae of Australian marsupials.
So we find that the Boopidae, Gyropidae, and Trimenoponidae occur upon marsupials, and upon South American Hystricomorph rodents, a group of which Matthew writes (p. 232):--"No hypothesis satisfactorily explains the accepted relationship and distribution." There is no evidence that these parasites have ever existed on other mammals in more northerly lands, and it seems most probable that they would have left residuals here and there if such had been the case. So here again the greater probability lies with Antarctic connection between South America and Australia. It may be that at a later stage Australia was cut off from Antarctica, and Africa joined, allowing of a migration of Hystricomorphs, but this raises a very complex question, bristling with difficulties, with which we are not here concerned.
The last instance I wish to bring forward is that of the distribution of crayfishes, also treated by Matthew (loc. cit., pp. 301-303). Matthew discusses the views of Ortmann (1902), and, after pointing out that the Holarctic crayfishes form a family distinct from the southern forms, which have a curious distribution in South America, New Zealand, Australia, and Madagascar, writes:--
"The Australian and South American crayfish I should regard as derived from the north, by way of the existing or slightly submerged land bridges, at a time when the northern crayfish were much more primitive than now and when, for reasons which I do not venture to suggest, the tropics were a more favourable environment than now. The northern crayfish have since evolved into Potamobius and Cambarus, the southern specialized into the more divergent Parastacus of South America, Cheraps and Engaeus and Astacopsis of Australia and Tasmania, Paranephrops of New Zealand . . . . and Astacoides of Madagascar.
"Of these southern genera, Astacoides is the nearest to the northern types. This is to be expected, if the southern genera are remnants of a cosmopolitan distribution derived by dispersal from the north; for the Malagasy genus would be a derivative from Ethiopian crayfish, which would be less remote from the north, and would be correspondingly more advanced than in South America and Australia."
Geoffrey Smith (1909, p. 214) refers to the distribution of crayfishes as follows:--
"It seems reasonable to suppose that the two families of crayfishes characteristic respectively of the northern and southern hemispheres have been independently derived from marine ancestors, which have subsequently become extinct. Their complete absence in the tropics is striking, and Huxley drew attention to the fact that it is exactly in those regions where the crayfishes are absent that the other large fresh-water Malacostraca are particularly well developed, and vice versa. Thus the large fresh-water prawns are typically circumtropical in distribution, while the South African rivers abound with River-crabs, which, in general, are found wherever crayfishes do not occur."
Later (loc. cit., p. 217) he writes of the general distribution of fresh-water Crustacea that it--"in the temperate southern hemisphere affords strong evidence in favour of the view that the chief land-masses of this hemisphere, which are at present separated by such vast stretches of deep ocean, were at no very remote epoch connected in such a way as to permit of an intermixture of the temperate fauna of New Zealand, Australia, and South America. While this connexion existed, a certain number of forms characteristic of the northern hemisphere, which had worked through the tropics by means of the Andes, were enabled to reach temperate Australia and New Zealand. The existence of a coast-line connecting the various isolated parts of the southern hemisphere would, of course, also account for the community which exists between their littoral marine fauna. [[p. 261]] It is impossible to enter here into the nature of this land-connexion which is becoming more and more a necessary hypothesis for the student of geographical distribution, whatever group of animals he may choose, but it may be remarked that the connexion was probably by means of rays of land passing up from an Antarctic continent to join the southernmost projections of Tierra del Fuego, Tasmania, and New Zealand."
He summarises these arguments again in his study of the Australian crayfishes (1912, p. 148):--
"The Astacidae (= Potamobiidae) and Parastacidae, the one family occurring in the northern Hemisphere, the other in the Southern, are therefore separated by important characters, and it is very probable that they have been independently evolved from marine lobster-like ancestors which already differed in these characters before they took to a freshwater life.
"The occurrence of Parastacidae in Australia, New Zealand, and South America, with an aberrant genus (Astacoides) in Madagascar--that is to say, in countries which are now separated by wide stretches of ocean--is a striking fact in geographical distribution, but it does not stand alone, the distribution of many freshwater fish, crustacea, molluscs, etc., having a similar character in the Southern Hemisphere.
"These facts, taken in conjunction with geological evidence, have led many naturalists to assume a much greater extension of the Antarctic Continent in past times which is supposed to have been connected with South America, Australia, and New Zealand, and possibly, at a very remote period with Madagascar, thus permitting the migration of land and freshwater animals to and from those countries. In the case of the Parastacidae the only alternative theory is that the South American, Australian, and New Zealand genera have been independently derived from some common marine ancestor."
This is a case in which parasites can be used to aid us. The four southern groups of crayfishes all carry ectoparasitic Temnocephaloids, a group generally associated with the Monogenetic Trematodes, though differing from these in certain important features. They are confined to fresh water, and are parasitic on the following hosts other than crayfish:--tortoises (Brazil), shrimps (Argentine), mollusc (Brazil), crab (Matto Grosso); shrimps and an Isopod (Australia); Crustacea (Java to Philippines). In addition, one species has apparently succeeded in reaching the northern crayfishes at their southern limit, Temnocephala mexicana being recorded from Cambarus digneti of Mexico.
Unless we make the assumption, for which there is no evidence, that Temnocephaloids were once found upon the Potamobiidae, but have become extinct upon all but Cambarus digneti, their distribution appears to prove quite conclusively the wide divergence between the northern and southern groups, which may have been due to a separate origin from hypothetical marine ancestors, as suggested by Smith, but which seems more likely to be due to the fact that, though having a common ancestry, they have been separated for a very long period of time, and the Parastacidae are simply less highly differentiated off-shoots of primitive Holarctic Potamobiids. From the greater variety of hosts upon which they are found in South America, it would seem that the Temnocephaloidea were evolved there, becoming parasitic upon the ancestors of Parastacus, and were carried with the migrating Crustacea to Antarctica, New Zealand, Australia, and Madagascar (perhaps by way of the Moluccas and Seychelles, as has been suggested for many other animals).
The weight of evidence is entirely against the separate colonisation from the north of New Zealand and Australia, and from Africa of Madagascar, which Matthew suggests. If crayfish had ever existed in Africa, they must have had Temnocephaloid parasites, since the Malagasy genus Astacoides has them, if it [[p. 262]] be presumed that the latter were derived from the former. It would follow that the Holarctic crayfish must have had these parasites. If so, where are they now? It is too much to ask us to believe that they have become extinct in the northern temperate zone when we find them so widely spread and holding their own in the southern. There is no evidence that crayfish have ever existed in the tropical belt, and the fact that their place is filled there by other creatures, such as freshwater crabs, and giant prawns, seems to indicate positively their non-existence at any time. The real explanation of the southern crayfishes seems to be that put forward by Geoffrey Smith (1909) that the Andes offered the only temperate path by which creatures of the northern zone could pass the tropics and enter the southern zone. This would account for the fact that many groups of animals which reached South America from the north have failed to reach either Africa or Australia.
We may conclude, then:--
1. That Matthew's hypothesis of Holarctic dispersal holds for man, and most, though not all, of the Tertiary mammalia, but that it does not hold for Mesozoic dispersal, nor for such groups as crayfishes and Hylid frogs.
2. That Australia has been isolated from the northern land masses since the late Mesozoic, and that those portions of its fauna which had to depend upon land migration have come in from the south.
3. That the Antarctic continent had, during the greater part of the Mesozoic and the early Tertiary a temperate climate, and a rain-forest flora, and was therefore habitable by animals.
4. That there are geographical indications of former connections between this continent and South America, New Zealand, Australia, and Africa, which, taken in conjunction with the evidence from faunal distribution, render it probable that land migration has from time to time been possible along such connections. No other hypothesis is adequate to explain some cases of distribution.
5. That evidence afforded by parasites strengthens that from faunal distribution, and minimises the suggestion of multiple and convergent origins.
6. That the Andes have always formed the only temperate pathway through the tropics.
7. That the Australian marsupials, together with the Parastacid crayfishes, the Leptodactylid and Hylid frogs, and a host of other forms, vertebrate and invertebrate, have reached Australia from South America by way of Antarctica.
Literature Cited.
BARBOUR, T. (1912).--A Contribution to the Zoogeography
of the East Indian Islands. Mem. Mus. Comp. Zool. Harvard, xliv.,
No. 1, pp. 1-168.
BENHAM, W. B. (1909).--Report on Oligochaeta
of the Subantarctic Islands of New Zealand. The Subantarctic Islands of
New Zealand, pp. 251-294. Wellington, 1909.
CHEESEMAN, T. F. (1909).--On the Systematic Botany of
the Islands to the South of New Zealand. Loc. cit. supra, pp.
389-471.
DAVID and PRIESTLEY (1914).--British Antarctic Expedition,
1907-9. Geology, Vol. i. London, 1914, Heinemann.
DUNN, E. R. (1923).--The Geographical Distribution of
Amphibians. Amer. Nat., lvii., No. 649, pp. 129-136.
FERRIS, G. F. (1922).--The Mallophagan Family Trimenoponidae.
Parasitology, xiv., pp. 75-86.
HARRISON, L. (1912).--The Taxonomic Value of Certain
Parasites. Abstract in Ann. Report. Syd. Univ. Sci. Soc., 1911-1912. JOHNSTON, S. J. (1913).--Trematode Parasites and the
Relationships and Distribution of their Hosts. Report. Aust. Ass.
Adv. Sci., xiv., pp. 272-278. KELLOGG, V. L. (1914).--Ectoparasites of Mammals. Amer.
Nat., xlviii., pp. 257-279.
LONGMAN, H. A. (1924).--The Zoogeography of Marsupials.
Mem. Queensland Mus., viii., pp. 1-15.
LONNBERG, E. (1921).--A second Contribution to the Mammalogy
of Ecuador with some Remarks on Caenolestes. Ark. Zool., xiv.,
No. 3, pp. 1-104.
MATTHEW, W. D. (1915).--Climate and Evolution. Ann.
New York Acad. Sci., xxiv., pp. 171-318.
METCALF, M. M. (1921).--Upon an Important Method of Studying
Problems of Relationship and of Geographical Distribution. Proc. Nat.
Acad. Sci., vi. NOBLE, G. K. (1922).--The Phylogeny of the Salientia.
Bull. Amer. Mus. Nat. Hist., xlvi., pp. 1-87. ORTMANN (1902).--Geographical Distribution of Fresh-water
Decapods and its Bearing upon Ancient Geography. Proc. Amer. Phil.
Soc., xli., pp. 267-400.
SMITH, G. (1909).--Crustacea, in the Cambridge
Natural History, Vol. 4. London, Macmillan. STILES and HASSALL (1908).--Index-catalogue of Medical
and Veterinary Zoology. Trematoda. Washington, 1908.
[[p. 263]]_____ (1914).--The Mallophaga
as a possible clue to Bird Phylogeny. Australian Zoologist,
i., pp. 7-11.
_____ (1915).--Mallophaga from Apteryx, and their Significance.
Parasitology, viii., pp. 88-100.
_____ (1915a).--The Relation of the Phylogeny of the Parasite to that
of the Host. Report. Brit. Ass. Adv. Sci., 1915, pp. 476-7 (Abstract).
_____ (1916).--Bird-parasites and Bird-phylogeny. Ibis, April,
1916, pp. 254-263.
_____ (1922).--The Mallophagan Family Trimenoponidae. Australian
Zoologist, ii., pp. 154-158.
_____ (1914).--Australian Trematodes and Cestodes; a Study in Zoogeography.
Medical Journ. Aust., Sept. 12, 1914 (Abstract in Report
Brit. Ass. Adv. Sci., 1914, p. 424).
_____ (1923).--The Opalinid Ciliate Infusorians. Bulletin 120, Smithsonian
Institution.
_____ (1923a).--The Origin and Distribution of the Anura. Amer. Nat.,
lvii., No. 652, pp. 385-411.
_____ (1924).--Contributions to the Herpetology of the Belgian Congo,
etc. Bull. Amer. Mus. Nat. Hist., xlix., pp. 147-347.
_____ (1912).--The Freshwater Crayfishes of Australia. Proc. Zool.
Soc. Lond., 1912, pp. 144-171.
_________________________
Notes Appearing in the Original Work
1. Italics mine. [[on
p. 248]]
2. Italics mine. [[on p. 248]]
3. Italics mine. [[on p. 252]]
4. Cystignathidae = Leptodactylidae.
[[on p. 256]]
5. Italics mine. [[on p. 256]]
6. Italics mine. [[on p. 256]]
7. Italics mine. [[on p. 256]]
*
*
*
*
*