
http://people.wku.edu/charles.smith/biogeog/DOBZ1950.htm
Evolution
in the Tropics
by Theodosius Dobzhansky (1950)
Editor Charles H. Smith's Note: Original pagination
indicated within double brackets. My thanks to American Scientist
for permitting the reprint. Citation: American Scientist 38 (1950):
209-221.
[[p. 209]] Becoming
acquainted with tropical nature is, before all else, a great esthetic
experience. Plants and animals of temperate lands seem to us somehow easy
to live with, and this is not only because many of them are long familiar.
Their style is for the most part subdued, delicate, often almost inhibited.
Many of them are subtly beautiful; others are plain; few are flamboyant.
In contrast, tropical life seems to have flung all restraints to the winds.
It is exuberant, luxurious, flashy, often even gaudy, full of daring and
abandon, but first and foremost enormously tense and powerful. Watching
the curved, arched, contorted, spirally wound, and triumphantly vertical
stems and trunks of trees and lianas in forests of Rio Negro and the Amazon,
it often occurred to me that modern art has missed a most bountiful source
of inspiration. The variety of lines and forms in tropical forests surely
exceeds what all surrealists together have been able to dream of, and
many of these lines and forms are endowed with dynamism and with biological
meaningfulness that are lacking, so far as I am able to perceive, in the
creations exhibited in museums of modern art.
Tropical rainforest impresses even a casual observer by the enormity of the mass of protoplasm arising from its soil. The foliage of the trees makes a green canopy high above the ground. Lianas, epiphytes, relatively scarce undergrowth of low trees and shrubs, and, finally, many fungi and algae form several layers of vegetational cover. Of course, tropical lands are not all overgrown with impenetrable forests and not all teeming with strange-looking beasts. One of the most perfect deserts in the world lies between the equator and the Tropic of Capricorn, in Peru and northern Chile. Large areas of the Amazon and Orinoco watersheds, both south and north of the equator, are savannas, some of them curiously akin to southern Arizona and Sonora in type of landscape. But regardless of the mass of living matter per unit area, tropical life is impressive in its endless variety and exuberance.
Since the animals and plants which exist in the world are products of the evolutionary development of living matter, any differences between tropical and temperate organisms must be the outcome of differences in evolutionary patterns. What causes have brought about [[p. 210]] the greater richness and variety of the tropical faunas and floras, compared to faunas and floras of temperate and, especially, of cold lands? How does life in tropical environments influence the evolutionary potentialities of the inhabitants? Should the tropical zone be regarded as an evolutionary cradle of new types of organization which sends out migrants to colonize the extratropical world? Or do the tropics serve as sanctuary for evolutionary old age where organisms that were widespread in the geological past survive as relics? These and related problems have never been approached from the standpoint of modern conceptions of the mechanism of evolutionary process. Temperate faunas and floras, and species domesticated by or associated with man, have supplied, up to now, practically all the material for studies on population genetics and genetical ecology.
Classical theories of evolution fall into two broad groups. Some assume that evolutionary changes are autogenetic, i.e., directed somehow from within the organism. Others look for environmental agencies that bring forth evolutionary changes. Although two eminent French biologists, Cuénot and Vandel, have recently espoused autogenesis, autogenetic theories have so far proved sterile as guides in scientific inquiry. Ascribing arbitrary powers to imaginary forces with fancy names like "perfecting urge," "combining ability," "telefinalism," etc., does not go beyond circular reasoning.
Environmentalist theories stem from Lamarck and from Darwin. Lamarck and psycholamarckists saw in exertion to master the environment the principal source of change in animals. Induction of changes in the body and the germ cells by direct action of physical agencies is the basis of mechanolamarckism. The organism is molded by external factors. A blend of psycholamarckist and mechanolamarckist notions, the latter borrowed chiefly from Herbert Spencer, has been offered by Lysenko as "progressive," "Michurinist," and "Marxist" biology. Advances of genetics have made Lamarckist theories untenable. There is not only no experimental verification of the basic assumptions of Lamarckism, but the known facts about the mechanics of transmission of heredity make these assumptions, to say the least, far-fetched.
Important developments and many changes have taken place in Darwinism since the publication of The Origin of Species in 1859. The essentials of the modern view are that the mutation process furnishes the raw materials of evolution; that the sexual process, of which Mendelian segregation is a corollary, produces countless gene patterns; that the possessors of some gene patterns have greater fitness than the possessors of other patterns, in available environments; that natural selection increases the frequency of the superior, and fails to perpetuate the adaptively inferior, gene patterns; and that groups of gene combinations of proved adaptive worth become segregated into closed genetic systems called species.
The role of environment in evolution is more subtle than was realized in the past. The organism does not suffer passively changes produced [[p. 211]] by external agents. In the production of mutations, environment acts as a trigger mechanism, but it is, of course, decisive in natural selection. However, natural selection does not "change" the organism; it merely provides the opportunity for the organism to react to changes in the environment by adaptive transformations. The reactions may or may
Fig. 1. Interior of equatorial rainforest near Belem, Brazil. The "terra firme" association. (Courtesy of Mr. Otto Penner, of the Instituto Agronomico do Norte, Belem do Pará.)
[[p. 212]] not occur, depending upon the availability of genetic materials supplied by the mutation and recombination processes.
Diversity of Species in the Tropics
Gause pointed out in 1934 that two or more species with similar ways of life can not coexist indefinitely in the same habitat, because one of them will inevitably prove more efficient than the others and will crowd out and eliminate its competitors. This "Gause principle" is a fruitful working hypothesis in studies on evolutionary patterns in tropical and temperate climates. The diversity of organisms which live in a given territory is a function of the variety of available habitats. The richer and more diversified the environment becomes, the greater should be the multiformity of the inhabitants. And vice versa: diversity of the inhabitants signifies that the environment is rich in adaptive opportunities.
Now, the greater diversity of living beings found in the tropical compared to the temperate and cold zones is the outstanding difference which strikes the observer. This is most apparent when tropical and temperate forests are compared. In temperate and cold countries, the forest which grows on a given type of terrain usually consists of masses of individuals of a few, or even of a single, species of tree, with only an admixture of some less common tree species and a limited assortment of shrubs and grasses in the undergrowth. The vernacular as well as the scientific designation of the temperate forest associations usually refers to these dominant species ("pine forest," "oak woodland," etc.). The forests of northern plains may, in fact, be monotonously uniform. In the forested belt of western Siberia one may ride for hundreds of miles through birch forests interrupted only by some meadows and bogs. Mountain forests are usually more diversified than those of the plains. Yet in the splendid forest of the Transition Zone of Sierra Nevada of California one rarely finds more than half a dozen tree species growing together. By contrast tropical forests, even those growing on so perfect a plain as that stretching on either side of the Amazon, contain a multitude of species, often with no single species being clearly dominant. Dr. G. Black, of the Instituto Agronomico do Norte in Belem, Brazil, made, in cooperation with Dr. Pavan and the writer, counts of individuals and species of trees 10 or more centimeters in diameter at chest height on one-hectare plots (100 x 100 meters) near Belem. On such a plot in a periodically inundated (igapó) forest 60 species were found among 564 trees. The numbers of species represented by various numbers of individuals on this plot were as follows:
Individuals
1 2 3
4 5 6 7
8 9
Species
22 9 7 2
2 2 4 2
1
Individuals 10 14 15 16
21 29 33 41 241
Species
1
1 1 1
1 1 1
1 1
The commonest species was the assaí palm (Euterpe oleracea), of [[p. 213]] which 241 individuals were found in the hectare plot, but as many as 22 species were represented by single individuals. On a plot of similar size, only a mile away but on higher ground (terra firma), 87 species were found among 423 trees. The numbers of individuals per species were as follows:
Individuals
1 2 3
4 5 6 7
9 12 17 20 25
37 49
Species
33 15 15 3 4
2 3 4 1 2
1 1 2
1
Here the commonest species was represented by only 49 individuals and as many as 33 species were found as single individuals. This high frequency of species represented by single individuals means that if
Fig. 2. The wet ground ("igapo") forest near Belem, Brazil. (Courtesy of Mr. Otto Penner, of the Instituto Agronomico do Norte, Belem do Pará.)
we had studied other hectare plots contiguous to the one actually examined, many new species would have been found. It is probable that each of the plant associations which we have sampled in the vicinity of Belem contains many more than 100 tree species, and, incidentally, only a few species occur in both associations. Similar results were obtained by Davis and Richards in the forests of British Guiana and by Beard in Trinidad.
The numbers of breeding species of birds recorded in the literature for territories in various latitudes, from arctic North America to equatorial Brazil, are as follows (data kindly supplied by Dr. E. Mayr):
Territory Number
of Species Authority
Greenland 56 F.
Salomonsen
Peninsular
Labrador 81 H.
S. Peters
[[p.
214]] Newfoundland 118 H.
S. Peters
New
York 195 K.
C. Parkes
Florida 143 S.
A. Grimes
Guatemala 469 L.
Griscom
Panama 1100 L.
Griscom
Colombia 1395 R.
M. de Schauensee
Venezuela 1148 W.
H. Phelps, Jr.
Lower
Amazonia 738 L.
Griscom
The numbers of recorded species of snakes are as follows (data obtained through the courtesy of Dr. C. M. Bogert):
Territory Number of Species AuthorityCanada 22 Mills
United States 126 Steineger and Barbour
Mexico 293 Taylor and Smith
Brazil 210 Amaral
The progressive increase in diversity of species from the Arctic toward the equator is apparent in general, even though there are irregularities in this increase, resulting from such factors as how the different territories compare in size, how uniform or varied they are ecologically and topographically, and how intensively their faunas have been studied. There can be no doubt that, for most groups of organisms, tropical environments support a greater diversity of species than do temperate- or cold-zone environments.
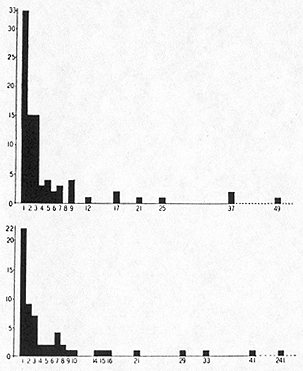
Fig. 3. Numbers of species of trees (ordinates) represented by different numbers of individuals (abscissae) on 1-hectare (100 x 100 meters) plots of equatorial rainforest near Belem, Brazil. "Terra firme" association (above) and "igapo" association (below). Only trees 10 or more cm. at chest height counted. (Data of Black, Dobzhansky, and Pavan.)
In order to survive and to leave progeny, every organism must be adapted to its physical and biotic environments. The former includes temperature, rainfall, soil, and other physical variables, while the latter is composed of all the organisms that live in the same neighborhood. A diversified biotic environment influences the evolutionary patterns of the inhabitants in several ways. The greater the diversity of inhabitants in a territory, the more adaptive opportunities exist in it. A tropical forest with its [[p. 215]] numerous tree species supports many species of insects, each feeding on a single or on several species of plants. On the other hand, the greater the number of competing species in a territory, the fewer become the habitats open for occupancy by each of these species. In the absence of competition a species tends to fill all the habitats that it can make use of; abundant opportunity favors adaptive versatility. When competing species are present, each of them is forced to withdraw to those habitats for which it is best adapted and in which it has a net advantage in survival. The presence of many competitors, in biological evolution as well as in human affairs, can be met most successfully by specialization. The diversity of habitats and the diversity of inhabitants which are so characteristic of tropical environments are conflicting forces, the interaction of which will determine the evolutionary fates of tropical organisms.
Chromosomal Polymorphism in Drosophila
Adaptive versatility is most easily attained by a species' becoming adaptively polymorphic, i.e., consisting of two or more types, each possessing high fitness in a certain range of environments. One of the most highly polymorphic species in existence is Homo sapiens, and it is this diversity of human nature which has engendered cultural growth and has permitted man to draw his existence from all sorts of environments all over the world. Now, the adaptive polymorphism of the human species is conditioned, on the cultural level, chiefly by the ability to become trained and educated to perform different activities. In species other than man, adaptive polymorphism is attained chiefly through genetic diversification--formation of a group of genetically different types with different habitat preferences.
In Drosophila flies, organisms best suited for this type of study, adaptive polymorphism takes its chief form in diversification of chromosome structure. In some American and European species of these flies, natural populations are mixtures of several interbreeding chromosome types which differ in so-called inverted sections. These chromosomal types have been shown, both observationally and experimentally, to have different environmental optima. The situation in tropical species has been studied in various bioclimatic regions in Brazil by a group consisting of Drs. A. Brito da Cunha, A. Dreyfus, C. Pavan, and E. N. Pereira of the University of São Paulo, A. G. L. Cavalcanti and C. Malogolowkin of the University of Rio de Janeiro, A. R. Cordeiro of the University of Rio Grande do Sul, M. Wedel of the University of Buenos Aires, H. Burla of the University of Zürich, N. P. Dobzhansky, and the writer. During the school year 1948-1949, the work of this group was supported by grants from the University of São Paulo, the Rockefeller Foundation, and the Carnegie Institution of Washington.
The commonest species in Brazil is Drosophila willistoni. It is also the adaptively most versatile species, since it has been found in every one of the 35 localities in various bioclimatic regions of Brazil in which collection was made. Significantly enough, this species, taken as a whole, [[p. 216]] shows not only the greatest chromosomal polymorphism among Brazilian species but the greatest so far known anywhere. The species Drosophila nebulosa and Drosophila paulistorum are also very common, but somewhat more specialized than Drosophila willistoni. Drosophila nebulosa is at its best in the savanna environments where dry seasons alternate with rainy ones, and Drosophila paulistorum, conversely, in superhumid tropical climates. The species are rich in chromosomal polymorphism, but not so rich as Drosophila willistoni in this respect. We have also examined some less common and biotically more specialized species, all of which showed much less or no chromosomal polymorphism.
The comparison of chromosomal polymorphism in Drosophila willistoni in different bioclimatic zones of Brazil proved to be even more interesting. The greatest diversity is found in those parts of the valleys of the Amazon and its tributaries where Drosophila willistoni is the dominant species. The exuberant rainforests and savannas of the Amazon basin are remarkable in the rich diversity of their floras and faunas; furthermore, the Amazon basin appears to be the geographical center of the distribution of the species under consideration, where it has captured the greatest variety of habitats. Yet, wherever in this region Drosophila willistoni surrenders its dominance to competing species, the chromosomal polymorphism is sharply reduced. This has been found to happen in the forested zone of the territory of Rio Branco, near Belem, in the state of Pará, and in the savanna of Marajó Island. In the first and the second of the regions named, Drosophila paulistorum, and in the third region, Drosophila tropicalis, reduce Drosophila willistoni to the status of a relatively rare species. In the peculiar desert-like region of northeastern Brazil, called "caatingas," Drosophila willistoni reaches its limit of environmental tolerance; Drosophila nebulosa seems to be the only species which still flourishes in this highly rigorous environment. The chromosomal variability of Drosophila willistoni is much reduced on the caatingas. It is also reduced in southern Brazil where Drosophila willistoni approaches the southern limit of its distribution, and is, presumably, losing its grip on the habitats.
Any organism which lives in a temperate or a cold climate is exposed at different periods of its life cycle or in different generations to sharply different environments. The evolutionary implications of nature's annually recurrent drama of life, death, and resurrection have not been sufficiently appreciated. In order to survive and reproduce, any species must be at least tolerably well adapted to every one of the environments which it regularly meets. No matter how favored a strain may be in summer, it will be eliminated if it is unable to survive winters, and vice versa. Faced with the need of being adapted to diverse environments, the organism may be unable to attain maximum efficiency in any one of them. Changeable environments put the highest premium on versatility rather than on perfection in adaptation.
[[p. 217]] Adaptive Versatility in the Tropics
The widespread opinion that seasonal changes are absent in the tropics is a misapprehension. Seasonal variations in temperature and in duration and intensity of sunlight are, of course, smaller in the tropical than in the extratropical zones. However, the limiting factor for life in the tropics is often water rather than temperature. Some tropical climates, for example those of the caatingas of northeastern Brazil, have variations of such an intensity in the availability of water that plants and animals pass through yearly cycles of dry and wet environments which entail biotic changes probably no less serious than those brought about by the alternation of winter and summer in temperate lands. Absence of drastic seasonal changes in tropical environments is evidently a relative matter. In Belem, at the opening of the Amazon Valley to the Atlantic Ocean, the mean temperature of the warmest month, 26.2° C., is only 1.3° C. higher than that of the coolest month, and the highest temperature ever recorded, 35.1° C., is only 16.6° C. higher than the all-time low, which was 18.5° C. The wettest month has 458 mm. of precipitation, and the driest has 86 mm., which is still sufficient to prevent the vegetation from suffering from drought.
It might seem that the inhabitants of the relatively invariant tropical climates should be free from the necessity of being genetically adapted to a multitude of environments, and hence that evolution in the tropics would tend toward perfection and specialization, rather than to adaptive versatility. This is not the case, however. The climate of Espirito Santo Island, in the tropical Pacific, is seasonally one of the most constant in the world as far as temperature and humidity are concerned. Nevertheless, Baker and Harrison found that native plants have definite flowering and fruiting seasons, and animals have cycles of breeding activity in this climate. Our observations in Brazil show that populations of Drosophila flies undergo expansions or contractions from month to month, as well as changes in the relative abundance of different species. The magnitude and speed of these changes are quite comparable to those which occur in California, for example, or in the eastern part of the United States. Such pulsations have been observed even in the rainforests of Belem, about 1½° latitude south of the equator. They are caused mainly by seasonal variations in the availability of different kinds of fruits which are preferred by different species of Drosophila. Despite the apparent climatic uniformity, the biotic environment of tropical rainforests is by no means constant in time.
This writer observed several years ago that certain populations of the fly Drosophila pseudoobscura which live in the mountains of California undergo seasonal changes in the relative frequencies of chromosomal types. Dubinin and Sidorov found similar changes in the Russian Drosophila funebris. What happens is that some of the chromosomal types of these flies possess highest adaptive values in summer, and other types in winter or in spring environments. Natural selection augments the frequency of favorable types and reduces the frequency of [[p. 218]] unfavorable types. The populations thus react to changes in their environment by adaptive modifications. This is one of the rare occasions when evolutionary changes taking place in nature under the
Fig. 4. Giant trees and lianas in the "terra firme" rainforest near Belem. (Courtesy of Mr. Otto Penner, of the Instituto Agronomico do Norte, Belem do Pará.)
influence of natural selection can actually be observed in the process of happening. We may add that some of these changes have also been [[p. 219]] reproduced in laboratory experiments in which artificial populations of the species concerned were kept in special "population cages."
We have observed populations of Drosophila willistoni in three localities in southern Brazil for approximately one year. One of the localities, situated in the coastal rainforests south of São Paulo, has a rather uniform superhumid tropical climate. Periodic sampling of these populations has disclosed alterations in the incidence of chromosomal types similar in character to those observed in California and in Russian fly species. Adaptive alterations which keep living species attuned to their changing environments occur in tropical as well as in temperate-zone organisms. This constant evolutionary turmoil, so to speak, precludes evolutionary stagnation and rigidity of the adaptive structure of tropical and of temperate species equally.
It is nevertheless true that tropical environments are more constant than temperate ones, in a geological sense. Major portions of the present temperate and cold zones of the globe underwent drastic climatic and biotic changes owing to the Pleistocene glaciation. The present floras and faunas of the territories that were covered by Pleistocene ice are composed almost entirely of newcomers. The territories adjacent to the glaciated areas have passed through more or less radical climatic upheavals. Although the bioclimatic history of the tropical continents is still very little known, it is fair to say that their environments suffered less change.
The repeated expansions and contractions of the continental ice sheets, and the alternation of arid and pluvial climates in broad belts of land bordering on the ice, made large territories rather suddenly (in the geological sense) available for occupation by species that could evolve the necessary adaptations in the shortest possible time. This has not simply increased the rates of evolution, but often has favored types of changes which can be characterized collectively as evolutionary opportunisms. Such changes have the effect of conferring on the organism a temporary adaptive advantage at the price of loss or limitation of evolutionary plasticity for further change. Here belong the various forms of deterioration of sexuality observed in so many species of temperate- and cold-zone floras. An apomictic or an asexual species, with an unbalanced chromosome number and a heterosis preserved by loss of normal meiotic behavior of the chromosomes, may be highly successsful for a time but its evolutionary possibilities in the future are more limited than those of sexual and cross-fertilizing relatives. Polyploidy is also a form of evolutionary opportunism in so far as it produces at least a temporary loss of genetic variability. Although some plant species native in the tropics have also become stranded in these evolutionary blind alleys, the incidence of such species is higher in and near the regions which were glaciated.
Evolutionary Importance of Biotic Environment
The contradictory epithets of "El Dorado" and "Green Hell," so often used in descriptions of tropical lands, really epitomize the two [[p. 220]] aspects of tropical environments. The process of adaptation for life in temperate and especially in cold zones consists, for man as well as for other organisms, primarily in coping with the physical environment and in securing food. Not so in the tropics. Here little protection against winter cold and inclement weather is needed. In the rainforests, the amount of moisture is sufficient at any time to prevent the inhabitants from suffering from desiccation. Relatively little effort is necessary for man to secure food, and it seems that the amount of food is less often a limiting factor for the growth of populations of tropical animals than it is in the extratropical zones. But the biological environment in the tropics is likely to be harsh and exacting. Man must beware that his blood does not become infected with malarial plasmodia, his intestines with hookworms, and his skin with a variety of parasites always ready to pounce on him and rob him of his vitality if not of life itself. The tremendous intensity of the competition for space among plants in tropical forests can be felt even by a casual observer. The apparent scarcity, concealment, and shyness of most tropical animals attest to the same fact of extremely keen competition among the inhabitants.
Now, the processes of natural selection which arise from encounters between living things and physical forces in their environment are different from those which stem from competition within a complex community of organisms. The struggle for existence in habitats in which harsh physical conditions are the limiting factors is likely to have a rather passive character as far as the organism is concerned. Physical factors, such as excessive cold or drought, often destroy great masses of living beings, the destruction being largely fortuitous with respect to the individual traits of the victims and the survivors, except for traits directly involved in resistance to the particular factors. As pointed out by Schmalhausen, indiscriminate destruction is countered chiefly by development of increased fertility and acceleration of development and reproduction, and does not lead to important evolutionary advances. Physically harsh environments, such as arctic tundras or high alpine zones of mountain ranges, are inhabited by few species of organisms. The success of these species in colonizing such environments is due simply to the ability to withstand low temperatures or to develop and reproduce during the short growing season.
Where physical conditions are easy, interrelationships between competing and symbiotic species become the paramount adaptive problem. The fact that physically mild environments are as a rule inhabited by many species makes these interrelationships very complex. This is probably the case in most tropical communities. The effectiveness of natural selection is by no means proportional to the severity of the struggle for existence, as has so often been implied, especially by some early Darwinists. On the contrary, selection is most effective when, instead of more or less random destruction of masses of organisms, the survival and elimination acquire a differential character. Individuals that survive and reproduce are mostly those that possess combinations of [[p. 221]] traits which make them attuned to the manifold reciprocal dependences in the organic community. Natural selection becomes a creative process which may lead to emergence of new modes of life and of more advanced types of organization.
The role of environment in evolution may best be described by stating that the environment provides "challenges" to which the organisms "respond" by adaptive changes. The words "challenge" and "response" are borrowed from Arnold Toynbee's analysis of human cultural evolution, although not necessarily with the philosophical implications given to the terms by this author. Tropical environments provide more evolutionary challenges than do the environments of temperate and cold lands. Furthermore, the challenges of the latter arise largely from physical agencies, to which organisms respond by relatively simple physiological modifications and, often, by escaping into evolutionary blind alleys. The challenges of tropical environments stem chiefly from the intricate mutual relationships among the inhabitants. These challenges require creative responses, analogous to inventions on the human level. Such creative responses constitute progressive evolution.
REFERENCES
1. ANONYMOUS. Normais Climatologicas. Serviço da Meteorologia. Ministerio da Agricultura. Rio de Janeiro, 1941.
2. BAKER, J. R. The seasons in a tropical forest. Part 7. Jour. Linn. Soc. London 41, 248-258, 1947.
3. BEARD, J. S. The natural vegetation of Trinidad. Oxford Forestry Mem. 20, 1-155, 1946.
4. BLACK, G. A., DOBZHANSKY, TH., and PAVAN, C. Some attempts to estimate the species diversity and population density of trees in Brazilian forests. Bot. Gaz., 1950. (In press.)
5. DA CUNHA, A. B., BURLA, H., and DOBZHANSKY, TH. Adaptive chromosomal polymorphism in Drosophila willistoni. Evolution, 1950. (In press.)
6. DAVIS, T. A. V., and RICHARDS, P. W. The vegetation of Moraballi Creek, British Guiana. J. Ecol. 22, 106-155, 1934.
7. DOBZHANSKY, TH. Observations and experiments on natural selection in Drosophila. Proc. 8 Internat. Cong. Genetics, Hereditas Suppl., 210-224, 1949.
8. DOBZHANSKY, TH., and PAVAN, C. Local and seasonal variations in relative frequencies of species of Drosophila in Brazil. Jour. Animal. Ecol., 1950. (In press.)
9. DUBININ, N. P., and TINIAKOV, G. G. Inversion gradients and natural selection in ecological races of Drosophila funebris. Genetics 31, 537-545, 1946.
10. GAUSS, G. F. The struggle for existence. Baltimore, 1934.
11. LACK, D. Darwin's finches. Cambridge, 1947.
12. PATTERSON, J. T. The Drosophilidae of the Southwest. Univ. Texas Publ. 4313, 7-216, 1943.
13. SCHMALHAUSEN, I. I. Factors of evolution. Philadelphia, 1949.
14. STEBBINS, G. L. Variation and evolution in plants. New York, 1950. (In press.)
15. TOYNBEE, A. J. A study of history. New York and London, 1947.
16. VANDEL, A. L'homme et l'évolution. Paris, 1949.
17. VAVILOV, N. I. Studies on the origin of cultivated plants. Leningrad, 1926.
*
*
*
*
*