
http://people.wku.edu/charles.smith/biogeog/BISB1943.htm
Geographical Distribution of Fungi
by Guy R. Bisby (1943)
Editor Charles H. Smith's Note: Original pagination
indicated within double brackets. Reprinted with permission from the New
York Botanical Garden Press. Originally published in The Botanical
Review, Vol. 9, pp. 466-482, copyright 1943, The New York Botanical
Garden.
[[p. 466]] INTRODUCTION
During a sojourn of some years in Manitoba, I became impressed by the large number of genera and species of fungi there in an environment that I had supposed would be unfavorable for them. Diligent collecting, by a few others and myself, disclosed 2,625 kinds, including 98 myxomycetes. It seemed evident that more than 5,000 species must be present (6), though only some 1,200 phanerogams are known to grow there without cultivation, and a hundred of these are introduced weeds or escapes. Green plants, then these fungi, had arrived and established themselves during not more than 25,000 years since the last glacier retreated.
After some thought and analysis I concluded (5) that at least twice as many phanerogams as fungi had been described for the world, but that in any state or country the number of species of fungi is likely to exceed the number of species of phanerogams because of the greater average range of a fungus; that the smaller the area surveyed, the greater is the excess of fungi; that saprophytic fungi generally have wider distribution than parasitic; and that distribution of hosts and substrata, principally flowering plants or their remains, has more influence than climate upon distribution of fungi.
Those conclusions are fairly obvious, but it should be possible to extend them. I now realize that a number of Manitoban fungi were not good morphologic species; that I had given most of my attention to the fungi of temperate regions, and therefore underestimated the effect of climate.
The study of distribution may be approached from various angles, from that of the whole world and all time to that of today's dispersal over one acre. I happened to be present during the arrival of a few fungi in Manitoba (5, 8); those recognised as new were all parasites of cultivated plants; I knew of no sure means of ascertaining when other fungi had reached the Province.
KRAKATAU: DISPERSAL IN OPERATION
The volcanic eruption of 1883 left the islands of the Krakatau group probably sterile, at least as regards fungi. These islands [[p. 467]] are now covered with a rich vegetation which has been carefully analysed. During the 50 years following the eruption a total of 263 species of phanerogams and 61 of pteridophytes had been found in gradually increasing numbers. Not all were able to persist, but in 1934, 219 phanerogams and 52 pteridophytes were reported (20). Immigration of fungi and lichens was not followed through the years, but a survey in 1933-4 gave 310 species of fungi, including myxomycetes, in 150 genera (9). Only 13 species of lichens were recorded.
The number of fungi was greater than that of vascular plants and would of course have been greater still if every fungus had been found and identified. The 310 species were grouped as follows: Myxomycetes, 28; Phycomycetes, 4 (all Mucorales, 3 of them species of Mucor isolated from the soil); Discomycetes, 10; Pyrenomycetes, 56; Uredinales, 2; Hymenomycetes, 150 (78 Polyporaceae); Gasteromycetes, 9; Fungi Imperfecti, 51 (34 Hyphomycetes).
Krakatau is about 40 kilometres from both Java and Sumatra. Boedijn found that a tree trunk with mycelium might occasionally be washed ashore and later produce spores, but he considered that nearly every fungus had arrived by air; for 50 years spores had settled on the islands, though none could develop unless its host or substratum had arrived. Twelve species are described as new from the Krakatau group, but, as Boedijn states, it is very unlikely that any of these is really endemic, i.e., "evolved" during the last 50 years.
Considerable search for lichens yielded only 23 collections belonging to 13 species, all epiphytes. None was found on rocks or soil, none on any of the islands except Krakatau. Lichens constitute most of the flora of the exposed rocks of Antarctica (33), but evidently these organisms rarely managed the trip from Java, Sumatra or elsewhere to Krakatau.
AGENTS OF DISPERSAL
We are here concerned particularly with the distribution of fungi over wide areas; but any agency that permits their spread may be considered. Fungi are estimated to have appeared upon the earth a couple of hundred million years ago, so they have not had to hurry. Many species now utilize more than one means of dissemination, e.g., ascospores by air, conidia by contact.
[[p. 468]] Dispersal with the aid of water and air has been analysed by Dobbs (19) and by Mason (28). Craigie (14) has reviewed aerial dissemination of fungi pathogenic to higher plants.
Air. It is recognized that light, powder-like seeds afford more rapid transit than plumed or winged seeds (36). Pteridophytes, with spores only about 30-50 microns in diameter, travel farther and faster than seed plants. Most fungi have even smaller spores; a spherical spore 5 microns in diameter has a volume of only 65 cubic microns, in contrast to 65,000 for such a spore 50 microns wide.
It is obvious that a slight current of air will support most fungus spores, provided they can escape from their sporophores. Nature has provided for this (11, 25): most basidiospores are launched by forcible abjection, most ascospores by ejection, and many other fungi, e.g., dry-spread Moniliales (28), Gasteromycetes, rusts and smuts, have powdery spores to be picked up by a breeze. Furthermore, most fungi produce spores in vast numbers. Viable spores have been caught thousands of feet up in the air. However, a spore is not likely to get across an ocean, or to be still alive if it does.
Even though a fungus has no device for getting its spores into the air, or though they fall to the ground, the wind may pick them up with dust.
Animals. Many fungi, such as Phallales, Sphaeropsidales, Melanconiales and about one-fourth of the Moniliales (34), have slime spores (28) spread by contact--with an animal, for example--or by water. Many different animals disseminate spores in sundry ways.
Insects have many relations with fungi, ably summarized by Leach (27). They may gather spores, particularly slime-spores, accidentally; often they are attracted to a spore-mass by odor (e.g., Phallales, decomposing vegetation), taste (e.g., a sweet secretion accompanying the conidia of ergot and the pycniospores of rusts), sight (brightly colored spore masses), or by appeals to two or three of these or comparable senses. Spores are then carried away on or in the insect's body.
Birds, particularly migratory birds, may carry spores long distances.
Man and his transport agencies have disseminated many fungi over the world. We can frequently trace such dispersal of parasitic fungi, commonly spread with seeds or other parts of plants, [[p. 469]] but it is seldom possible to decide whether a saprophyte is native or introduced.
Water. Rivulets, streams and spattering rain spread fungi. Zoospores of Myxomycetes and Phycomycetes develop and propel themselves in water. Floating leaves, stems or fruits may transport fungi in fresh or even in sea water. Many spores will float. Hygroscopic movements may aid spore discharge.
Several agencies and factors may interact to determine the effective distribution of an organism. A living fungus spore may travel far, but it must land on the proper host or organic substratum, then develop more spores, despite all vicissitudes. Even then the species may not be able to persist. We know so little of the distribution of most fungi that we can seldom say when, whence, often not even whether, a fungus has arrived in any country.
EFFECT OF CLIMATE ON DISTRIBUTION
Climate is of primary importance in the distribution of vascular plants, the host or substratum in the distribution of fungi. In other words, climate affects fungi more indirectly, or at a stage once removed. Phanerogams cultivated over wide areas, e.g., corn, tomatoes, potatoes, have numerous parasites that attack them anywhere. Nevertheless, climate has a controlling effect on the distribution of many fungi, including various parasites of crop plants.
We are not surprised to find certain species of Mucor or Botrytis everywhere, of Peziza or Lycoperdon almost anywhere, of Puccinia or Cercospora wherever a host thrives. But the range of a fungus is often quite different from that of its hosts or substrata; climatic or other factors must then be involved in limiting its distribution.
Dry regions are characterized by xerophilous Gasteromycetes, Agaricaceae and other fungi, many of which are restricted further to warmer dry localities. Phymatotrichum omnivorum, the cause of Texas root-rot of cotton, is definitely known only in southwestern North America; it attacks hundreds of hosts but has not followed them northward nor into moist regions. A dry atmosphere of course limits the development of many leaf parasites and others, but may even favor some, such as certain powdery mildews.
Damp temperate regions, such as those of northern Europe and eastern North America, are generally taken to have the "standard" fungus flora, i.e., the one first and best known.
[[p. 470]] The moist tropics contain many fungi, a considerable proportion of which are found nowhere else. Most of the accompanying phanerogams are of course also limited to the tropics; but with some groups of fungi it is evidently the climate, not the host, that controls distribution. The Meliolaceae, for example, develop abundantly on diverse hosts in warmer regions; they are few and rare even on similar hosts in cooler areas.
A fungus in the tropics does not have to tide itself over a cold winter, though it generally has to be prepared for a dry spell. Erysiphaceae and various rusts in warm areas persist in their conidial stages.
Arctic regions seem to have high proportions of Pyrenomycetes, frequently without conidial stages, and perhaps more plurivorous than those in temperate countries (18).
Bose (10) reported that the phanerogams and a half dozen Polyporaceae of the Lokra Hills, India, are European in character. He rightly considered that the hosts were of primary importance in the distribution of these polypores; but I think we may say that climate also had an effect. He includes Polyporus sulphureus and P. squamosus, species with a wide host range in the North Temperate Zone, the former being known in Britain on various dicotyledonous and coniferous trees. I do not know how or when these two fungi reached the Lokra Hills (they occur also in Himalayan areas), but, since their potential host range is so wide, it would seem that climate helps to keep them there.
Temperature, precipitation, humidity, length of season and other climatic factors affect the distribution of fungi. It is not always easy to evaluate these factors. Claviceps purpurea, the cause of ergot, would seem to be limited by temperature to temperate regions; but it is possible that Cerebella, a fungus which grows on conidial (Sphacelia) stages in warm countries, may be more important than climate.
AGE AND AREA
Every student of distribution is in debt to Willis (36, 37) for his extensive compilations and analyses. A few data in this and subsequent sections may be interpreted as having a bearing on his hypothesis of Age and Area, i.e., that (with qualifications) the area occupied by a group of allied species depends chiefly upon or varies directly with their ages.
[[p. 471]] The exact geographic distribution is known for few of the 3,600 genera and 37,000 species of fungi accepted as "good" (1, 7). Nearly half (47.2%) of the genera of fungi are at present monotypic, as compared with 38.6% of the genera of vascular plants. Fungi show the "hollow curve" stressed by Willis, i.e., the decreasing number of genera having 1, 2, 3 . . . species. The figures for representative groups of fungi are given in Table I.
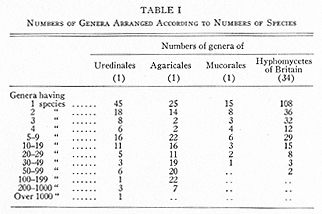
Willis finds, in general, that the more species per genus, the greater the area covered; the older the genus or species, the greater its area. An analysis for Uredinales is given below.
Rhizophagus is one of the very few fungi that can be traced through the geologic ages. It is now widely distributed on underground parts of Hepaticae, ferns and flowering plants; a fungus of similar morphology is found on fossil remains back to those of the Psilophytales, the earliest land plants (12). Despite its age and wide distribution, Rhizophagus is credited with only three species.
Myxomycetes are generally considered to be amongst the more primitive organisms. They are also very widely distributed, though there are relatively few species. Not much is known of the distribution of most other older groups, such as the Chytridiales.
Some fungi are "being evolved" today, i.e., attaining characters which lead mycologists to name them as new species or genera. [[p. 472]] Mutations are frequently recognized in the laboratory, sometimes in the field. Parasitic fungi are likely to adapt themselves, with or without morphologic changes, to a new phanerogam as, or soon after, it appears; saprophytic fungi, on the other hand, are unlikely to require any adaptation to the remains of a new plant.
DISTRIBUTION OF GENERA OF FUNGI
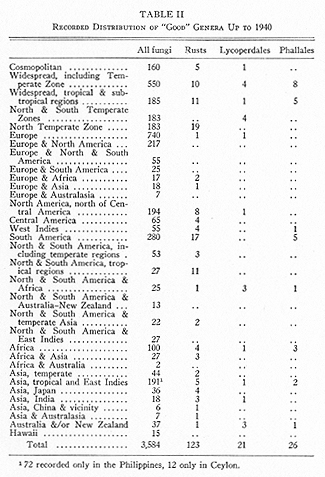
An analysis of reported distribution of genera of all fungi (accepted in (1)) is given in Tables II to IV. It will be understood (a) that many generic names have little or no real basis, or are recognised by one mycologist and not by another; (b) that the actual distribution of most genera is greater than now known; (c) that the figures frequently represent numbers and activity of mycologists rather than distribution: for example, 740 genera are "endemic" in Europe, only 194 in North America. This is approximately the ratio of European and North American taxonomists (237 and 50) listed in our dictionary (1), with allowance for the fact that European mycologists have described numerous North American genera. It is true that only a half dozen mycologists have devoted themselves exclusively to naming the fungi of South America where 280 endemic genera are reported; but the activity of Spegazzini and of taxonomists outside South America has made up for any shortage of workers. Africa, with only 100 genera supposedly endemic, is still rather dark--or conservative--mycologically.
Despite the uncertainties, it seems clear that the distribution of fungi differs from that of phanerogams. Considerable collecting in Ceylon and in Hawaii--islands noted for high percentages of endemics--has shown only 12 and 15 genera, respectively, of fungi not yet known elsewhere. The considerable number, 72 genera, known only in the Philippines is likely to be revised in time, though one would expect endemic fungi there. In areas such as Canada, with only a half dozen genera of fungi not known elsewhere, and Manitoba with no endemic genus of fungi or of vascular plants, the distribution of fungi is like that of phanerogams.
Table III gives an approximate summary of Table II. It indicates more clearly that many genera and therefore species of fungi are widely distributed.
[[p. 473]]
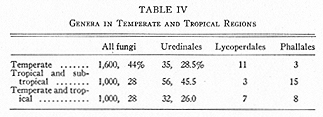
Another calculation gives the figures shown in Table IV, where the column for "all fungi" is influenced by the greater study given to fungi in temperate regions.
[[p. 474]]
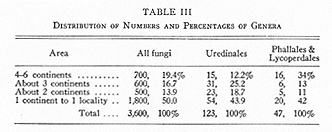
Table V gives an idea of the number of genera of fungi recorded from each of the continents. For "all fungi" the figures have been
reduced in an effort to discount dubious names and useless "segregates".
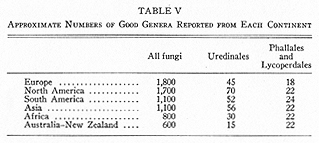
The figures for all fungi in Tables II-V leave one rather confused. We turn, therefore, to three orders with better known distribution: the Uredinales, all obligate parasites, and the Phallales and Lycoperdales, all saprophytes.
[[p. 475]] DISTRIBUTION OF UREDINALES
Distribution of rusts is dependent on that of their hosts--or even, for heteroceious rusts, on that of two dissimilar kinds of hosts. Nevertheless, since a species of rust is usually able to attack more than one species of host, the average range of a rust may be greater than that of a host.
It is somewhat surprising to note that only two species of rust, and only two collections of each, have been found on the Krakatau islands; for we know that many rusts, notoriously Puccinia graminis, travel far by means of air-borne urediospores.
About 60% of the rusts of North America are endemic; at least 40 species (4%) have been introduced by man (2). Temperate parts of that continent have the largest number of rusts; boreal parts have the fewest but show a higher percentage of Melampsoraceae and of microcyclic species.
An analysis based on Table I shows that genera with more than 200 species (Puccinia, Uromyces, Aecidium, Uredo) are all cosmopolitan, though the genus (Ravenelia sensu lat.) next in size is almost confined to Leguminosae and to warmer areas. The 45 monotypic genera are mostly restricted in distribution: North Temperate Zone, 2; Europe, 1; Europe and Africa, 2; Africa, 4; Java, 1; Philippines, 2; Japan, 3; India, 2; North America (Mexico north), 7; Central America, 4; West Indies, 4; South America, 12; Tropical America (North and South), 1. All this is in harmony with "Age and Area". One rather striking fact is that 28 (62%) of these monotypic genera are known only in the Americas, mostly in tropical or sub-tropical areas. This perhaps to some extent reflects the rather greater study of rusts in the western hemisphere than in tropical Africa or Asia; but it suggests that warmer American regions are or have been a main center of rust development. Genera with two and three species, though naturally rather more widely distributed than monotypic genera, are also to a considerable extent American. Hemileia, however, is predominantly African, and about half its species attack Rubiaceae.
Cummins (16) has made a thorough study of Prospodium. All 50 species, except one doubtful form of Uredo from Ceylon, known only from the type collection showing no telia, have been found in warmer American regions. Most of them are on Bignoniaceae, 22 genera of which are recorded as hosts; nine species are on [[p. 476]] Verbenaceae--one on Lantana, eight on Lippia. No other host family is known. The Bignoniaceae occur particularly in South America; the Verbenaceae, though also mostly limited to warmer countries, are not predominantly American. I conclude that hosts primarily, then climate, limit the distribution of Prospodium; that the genus and species obviously evolved in warmer America; that for some reason, possibly lack of time and opportunity, little or no spread to other countries and other related hosts has taken place.
It is noteworthy that only about 30 genera of Uredinales (four endemic) were recorded from Africa up to 1940 (Tables II-V); that only 15 were known in Australia and New Zealand, only one of which is endemic despite the high proportion of endemic higher plants which exist there as potential or actual hosts. New Zealand (17) has 53% of endemic species of rusts, rather less than the North American percentage, but, as might be expected, it has nearly as many introduced species (33 in New Zealand, 40 in North America).
I am inclined to look with suspicion upon a fungus which remains "known only from the type locality". There are some 35 rusts, exclusive of those in form genera, in this class in the United States and Canada. Uredinologists will doubtless seek explanations for such apparent rarity; perhaps some of these species are already eligible for a group of fungi hitherto unknown, at least to me, namely, those that have definitely disappeared from the earth since man described them.
It is well known that similar rusts occur in alpine regions in various parts of the world; for example, 22% of the rusts in alpine regions in Japan are found also in the Swiss Alps (23). I interpret this as primarily a host relationship, though climate is involved, as it is particularly in the shortening of life cycles at high altitudes.
The distribution of many rusts seems perplexing at the moment (e.g., Trillium rust (3)), but important facts regarding the distribution and evolution of parasitic fungi have come and will come from studies on Uredinales (26). Many of them are widespread, more than a third of the genera being known from three or more continents (Table III).
Crowell (15), in a comprehensive study of the distribution of Gymnosporangium, concludes that various factors limit many species to ranges less than that of their hosts, and considers this quite [[p. 477]] the reverse of my general finding (5). It still must be remembered, however, that most species of Gymnosporangium have more than one aecial or telial host. Take G. juvenescens, for example; its distribution seems to cover only about half that of its hosts--but Arthur records it on eleven aecial and three telial hosts. My thesis was that an average rust covers more range than an average host, but not necessarily more than a dozen of its hosts.
DISTRIBUTION OF GASTEROMYCETES
The conspicuous or even bizarre nature of many Gasteromycetes has resulted in exceptional knowledge of their distribution. Australia-New Zealand stand out by having about as many genera and as many endemic species as are to be found in any larger continental area. Primitive Gasteromycetes are found particularly in regions bordering the Pacific, indicating that these regions have been centers for development of the class (21). An analysis of genera of Phallales and Lycoperdales having one, two and three species suggests the same conclusion: about half such genera in each of the two orders are known near the Pacific. It is perhaps even more noteworthy that of eleven monotypic genera of Phallales, ten are restricted to warm and relatively small areas in America, Asia or Africa, the eleventh to New Zealand. Table IV shows that the genera of Phallales are predominantly tropical, of Lycoperdales temperate. Nearly half the genera in these orders are still known only within a single continent, in spite of the fact that man has introduced several Phallales to distant countries: Lysurus and Aseroe to Britain, for example; probably Colus ?javanicus to the United States (31). One may conclude that many Phallales had a rather limited distribution before man began to travel; in other words, that they are influenced by climate and probably by having had less time than many other fungi in which to travel by their own devices.
Other Gasteromycetes worthy of mention are Queletia with one species known from three collections in France, one in Britain, and a couple in successive years in one spot in Pennsylvania; and Podaxis with one species in dry regions within about 40° north and south of the equator (29). Incidentally, the dependence of knowledge of distribution upon that of taxonomy is well illustrated with Podaxis; before Miss Morse's study one might have tabulated a [[p. 478]] couple of dozen "species" of the genus, many of them recorded from only one locality.
Few saprophytic fungi in other groups can be so definitely recognized as having been introduced to an area. Many Gasteromycetes are widespread, but it is not surprising that some of these specialized "highest" fungi should have limited natural ranges. Once more we see that climate is important, even to obligate saprophytes.
DISTRIBUTION OF OTHER GROUPS
Myxomycetes are amongst the best known, most widely distributed and presumably oldest of fungal groups. We have noted that 28 species, 7% of all, are reported to have reached the Krakatau islands.
Little is known of the distribution of most of the simple, often aquatic, Phycomycetes; but some Peronosporales and Zygomycetes are cosmopolitan. During the last few years, Drechsler (22) has found a whole new family, the Zoopagaceae, belonging to the Entomophthorales; the species are probably widely distributed, but at present most of them are reported from only one or two American collections.
Ascomycetes contain many widespread fungi and many with unknown distribution. What can a student of plant geography do with a genus such as Dipodascus? It has two species, one described 50 years ago from Ecuador, and since then recognised twice, from Sweden and Colombia; the second species has but one record, from Canada (4). The Laboulbeniales seem to develop most abundantly in the tropics, though 150 species are said to be palaearctic, with more than 50 recorded from Poland (32).
The Hymenomycetes of the north temperate area are similar everywhere; some species, such as Armillaria mellea, extend over much of the world. In most groups, however, the species in tropical or south temperate regions are to a considerable extent distinct from those in the north temperate. It should be noted, however, that coprophilous agarics and other coprophilous fungi are almost as ubiquitous as their substratum; that the genera of Hymenomycetes in the south temperate (13) are about the same as those in the north, and that even in the torrid zone the majority of genera of Hymenomycetes are those of temperate regions.
Overholts, in an important contribution (30) to the knowledge of geographical distribution, finds that 43% of the American pileate [[p. 479]] Polyporaceae are known in Europe or other parts of the eastern hemisphere, but that many species are limited by climatic factors. He has been able to map a number of species, and to predict extensions to their known distribution in the United States and Canada. When accurate maps such as his become available for other groups of fungi, we can start to make more definite generalizations on distribution.
Many Fungi Imperfecti, particularly Hyphomycetes of the "mould" group such as those in the soil, are to be found anywhere.
DISTRIBUTION OF FUNGI PARASITIC ON CROP PLANTS
It is scarcely necessary to discuss in detail the distribution of crop diseases. Maps are now being published (24) showing the range of many. Man has been very active in assisting nature; for example, asparagus rust was enabled to establish itself from New Jersey to California in five years.
A few pathogens are worthy of mention because they seem to illustrate principles. Puccinia Antirrhini, the rust of snapdragons, is native to a few wild Scrophulariaceae in the mountains of California. Soon after Antirrhinum majus was introduced there it was attacked and proved to be a very congenial host; the rust spread on snapdragons throughout the United States and Canada, and now occurs over much of Europe and in Egypt, Palestine and South Africa.
The original host of Synchytrium endobioticum, the cause of wart disease of potatoes, is not known. Potatoes were in general culture in Europe for about 150 years (35) before the fungus was described on them in 1896. It then spread over northern Europe and reached Newfoundland and South Africa, but its late start permitted prevention of its spread over North America.
Spongospora subterranea, another parasite of the potato, has been known for a century. It has spread far. Considerable alarm was felt in North America after it was first found in Canada in 1913, but subsequent experience demonstrated that climate almost limits it to cool regions such as those near the United States-Canada boundary in the east and west. The fungus seems to be unimportant, except perhaps at high altitudes, in Asia, Africa and South America. Possibly a consideration of the effect of climate on this and other pathogenic fungi would permit some modification of the elaborate quarantine and inspection regulations imposed by most [[p. 480]] countries. Unfortunately, we do not yet know how much faith to place in climate. Can we be sure that Claviceps purpurea will not develop in the tropics, and only in Algeria and Morocco in Africa?
Cronartium ribicola has now spread over most north temperate regions where its hosts grow, Ribes and five-needle pines in association. The same is true of many another parasite of economic plants. In other words, the host is of primary importance in dissemination of parasites. It would be hard to stop nature and man in their efforts to spread pathogens, were it not that climatic and other factors are also important, and may hinder as well as favor spread.
RESTATEMENT
Comprehensive facts and principles regarding geographical distribution are scarcely possible until many more fungi can be mapped accurately. Taxonomy is badly in need of synthesis, so that mycologists everywhere can speak in the same terms. Much progress has, however, been made during the decade since I previously attempted to summarize the subject.
The morphological and physiological devices and processes enabling fungi to present their spores for dispersal are now seen much more clearly. Spores have been tracked through the upper air, across continents, and to new Krakatau. A more definite picture of the mycological flora of the tropics and south temperate is developing.
We conclude (7), after going over the fungi genus by genus, that less than 40,000 good species are known today, and many of these are "host-species" based on host plus morphology. Some 60% of all species are saprophytes. Perhaps 100,000 or even more species may be present on earth. There are about 200,000 known species of phanerogams, and I suppose about 100,000 still unknown, unless taxonomists turn more conservative and reduce their numbers.
Climate has a controlling effect on the distribution of many fungi. There are other factors, such as plant succession, amount of disturbance of the environment, light and length of season, the effects of which are difficult to evaluate. Study of the distribution of monotypic genera and of important parasites or other species which can be mapped in detail, with application of the principles of statistics, philosophy, and "Age and Area", seem likely to prove illuminating.
[[p. 481]] I now venture this revised summary to replace the one I made ten years ago:
1. Species of phanerogams acceptable today outnumber such species of fungi by about five to one. There are perhaps three times as many phanerogams as fungi on earth.
2. Nevertheless, species of fungi can be expected to outnumber species of phanerogams in any particular state or country. In other words, the average range of a fungus is the greater.
3. The smaller the area surveyed, the more the fungi predominate.
4. Though certain fungi were doubtless amongst the most primitive inhabitants of the earth, they have come to depend largely on phanerogams and their remains.
5. Saprophytic fungi generally have wider distribution than parasitic.
6. Distribution of hosts and substrata primarily controls distribution of fungi. A parasite is commonly able to attack more than one species of host, so that its range can be greater than that of any one of its hosts.
7. Climate has an important influence on many fungi. Even obligate saprophytes may be limited to certain climatic areas.
8. Other factors, including the nature and density of phanerogams, light, and particularly the activities of man, influence the distribution of many fungi.
LITERATURE CITED
1. AINSWORTH, G. C. and BISBY, G. R. A dictionary of
the fungi. [In press.]
2. ARTHUR, J. C. The plant rusts. 1929.
3. BARRUS, M. F. Trillium rust. Mycologia 20: 117-126. 1928.
4. BIGGS, R. Dipodascus uninucleatus. Mycologia 29:
34-44 [See also p. 619]. 1937.
5. BISBY, G. R. The distribution of fungi as compared with that of phanerogams.
Am. Jour. Bot. 20: 246-254. 1933.
6. _____ et al. The fungi of Manitoba and Saskatchewan. Nat.
Res. Council, Canada. 1938.
7. _____ and AINSWORTH, G. C. The numbers of fungi. Trans. Brit. Mycol.
Soc. 26: 16-19. 1943.
8. _____ and CONNERS, J. L. Plant diseases new to Manitoba. Sci. Agr.
8: 456-458. 1928.
9. BOEDIJN, K. B. The Mycetozoa, Fungi and Lichenes of the Krakatau group.
Bull. Jard. Bot. Buitenzorg, III. 16: 358-429. 1940.
10. BOSE, S. R. The distribution of some Polypores at our high altitudes.
Ann. Mycol. 23: 201. 1935.
11. BUTLER, A. H. R. Researches on fungi. Vols. I-VI. 1909-34.
12. BUTLER, E. J. The occurrence and systematic position of the vesicular-arbuscular
type of mycorrhizal fungi. Trans. Brit. Mycol. Soc. 22: 274-301. 1939.
[[p. 482]] 13. CLELAND, J. B. Toadstools
and mushrooms and other larger fungi of South Australia. 1934-5.
14. CRAIGIE, J. H. Aerial dissemination of plant pathogens. Proc. VI Pacific
Sci. Congr. 1939. 1941.
15. CROWELL, I. H. The geographical distribution of the genus Gymnosporangium.
Canad. Jour. Res. 18C: 469-488. 1940.
16. CUMMINS, G. B. The genus Prospodium (Uredinales). Lloydia
3: 1-78. 1940.
17. CUNNINGHAM, G. H. The rust fungi of New Zealand. 1931.
18. DEARNESS, J. Fungi. Report of the Canadian Arctic expedition 1913-18,
4C: 1-24. 1923.
19. DOBBS, C. G. On the primary dispersal and isolation of fungal spores.
New Phyt. 41: 63-69. 1942.|
20. DOCTERS VAN LEEUWEN, W. M. Krakatau, 1883 to 1933. Ann. Jard. Bot.
Buitenzorg, 46-7: 1-506. 1936.
21. DODGE, C. W. Contribución al conocimiento de la evolución
de los Gasteromicetos. Rev. Sudamer. Bot. 1: 18-20. 1934. [From Biol.
Abs. 10, No. 16,751.]
22. DRECHSLER, C. New Zoopagaceae capturing and consuming soil amoebae.
Mycologia 30: 137-157. 1938.
23. HIRATSUKA, N. A contribution to the knowledge of the rust-flora in
the alpine regions of high mountains in Japan. Mem. Tottori Agr. Coll.
3: 125-247. 1935.
24. Imperial Mycological Institute. Distribution maps of plant diseases.
1942-43.
25. INGOLD, C. T. Spore discharge in land plants. 1938.
26. JACKSON, H. S. Present evolutionary tendencies and the origin of life
cycles in the Uredinales. Mem. Torrey Bot. Club 18: 1-108. 1931.
27. LEACH, J. G. Insect transmission of plant diseases. 1940.
28. MASON, E. W. Annotated account of fungi received at the Imperial Mycological
Institute. List 2, Fasc. 3. 1937.
29. MORSE, E. E. A study of the genus Podaxis. Mycologia 25:
1-33. 1933.
30. OVERHOLTS, L. O. Geographical distribution of some American Polyporaceae.
Mycologia 31: 629-652. 1939.
31. SEAVER, F. J. A rare phalloid from the New York Botanical Garden.
Mycologia 23: 83-84. 1931.
32. SIEMASZKO, J. & W. Laboulbeniales polonici et palaearctici, II.
Polskie Pismo Entomol. 10: 149-188. 1931.
33. SIPLE, P. A., DODGE, C. W. and BAKER, G. E. The second Byrd Antarctic
Expedition--Botany. Ann. Missouri. Bot. Gard. 25: 467-718. 1938.
34. WAKEFIELD, E. M. and BISBY, G. R. List of Hyphomycetes recorded from
Britain. Trans. Brit. Mycol. Soc. 25: 49-126. 1941.
35. WEISS, F., ORTON, C. R. and HARTMAN, R. E. Investigations of potato
wart. U. S. Dept. Agr., Dept. Bull. 1156. 1923.
36. WILLIS, J. C. Age and area. 1922.
37. _____. The course of evolution. 1940.
*
*
*
*
*