The
Once and Future Wallace
by Charles H. Smith, Ph.D.
Abstract: Biogeography has always been
recognized as an interdisciplinary subject, but the relations involved
are often referred to particular traditions of research only, and not
to unifying principles that might cross such boundaries. Thus, historical
biogeographers tend to emphasize the way past geographic relationships
might influence species dispersal and divergence in a deterministic,
black box fashion that neglects the ecological roles of the actors,
whereas ecological biogeographers have often explained presence or absence
on the basis of conditions of the immediate environment that are poorly
linked to longer term, evolutionary, history. Through the use of the
concept of final causes an attempt is made here to steer thinking toward
population, community and ecosystem integration models that are less
divisive in this regard. Applications to the study of mass extinctions,
ten-year cycles in northern wildlife populations, and natural biogeographic
range change are briefly described. Keywords: environmental
evolution, dinosaur extinction, atmospheric oxygen, atmospheric evolution,
final causes, simulation, ubiquitous and nonubiquitous resources, Hutchinsonian
niche.
1. Introduction
It was apparently Jean-Paul Sartre who once commented that there must be a God, as otherwise all eventualities would be permissible. For those of a non-theistic bent, this notion may be re-stated to read that there is only one way to understand how things can not be permitted: through admitting the existing agency of countervailing forces. And in fact we do have what we have, and not innumerable other realities, because in one manner or another forces got in the way that prevented other things from happening. We know this; it is obvious, in fact. Nevertheless, it has proved very difficult over the history of scientific thought to identify exactly how such forces--at all levels of scale and intensity--have conspired to produce the kinds of irreversible change we witness. In the following essay I should like to consider briefly certain elements of this problem, especially those of a basic philosophical kind relating to notions of causation, and some of an exploratory nature bearing on the interrelation of organismal and environmental change.
2. Causes, and Teleological Explanation
The basic Aristotelian stance on causation--the four causes model--is the standard starting point for all discussions on the philosophy of ordered change. Aristotle organized his natural philosophy around the complementary notions of efficient, material, formal, and final (or end) causation. As one source indicates, however, the "concepts of material and formal causes are archaic and now have little significance outside aesthetics" (Taylor 1967, p. 56), with the result that scientific endeavor has mostly concerned itself with identifying efficient and final causes, with a good deal more emphasis having been put on the former than on the latter. Aristotelian thinking originally took rather the opposite position: final causes were linked to ultimate functions within nature while efficient causes were connected to the mechanics of sequences of events only. Aristotle's investment in final causes leads naturally to the notion of teleology, in which the events and structural results of a later time are viewed as to one degree or another preordained.
Teleological forms of explanation, and the concept of final causation altogether, have usually been regarded by modern science as antithetical a priori to dispassionate analysis, but perhaps there are elements of this way of thinking that should not be quite so quickly dismissed. It first should be noted that while it is true that most theistic explanations of reality rely on a first-causes-based teleology operationalizing Godly intervention, teleology per se need not invoke assumptions of the "interference" by miraculous forces. Actually, wherever conscious acts of will are invoked toward the end of producing a particular result, final causation seemingly remains a useful tool for contextualizing the process involved. The problem comes, however, in attempting to understand how final causes might operate where, apparently, no conscious will is involved.
This is a slippery slope, tempting the unwary to fall into various kinds of counterproductive thinking. One tendency is to accede to a form of pantheistic super-nature in which everything existing "knows its place" and acts accordingly--a view bereft of explanatory value. Or, one may be deluded into giving into a depressing brand of predestination and come to believe in absolute fates. But as with most extremes, there are more reasonable alternatives available to choose from.
Evolution is often viewed as tree-like in its emergence and expression; certainly, this is the usual metaphor attached to the process--continuing divergence of species lines-- we know as phylogenesis. But phylogenesis as we look back on it is not evolution, it is merely a recapitulation of the way the processes involved in evolution played out. It is how all those foci of information storage interacted with one another through the medium of environment, and how the environment itself was affected, that truly constitutes the process we term evolution. It is here where we have to face up to Sartre's dictum: in evolution we are not dealing with a structure produced through a simple, unimpeded random walk kind of causality, but instead one which is confronted at every turn by immediate, if sometimes temporary, impasses: in short, by countervailing forces.
The truth of the matter is that we have hardly begun to understand the degrees to which such forces affect the overall process, and at what scales, spatial and temporal. There seems no reason to doubt that all sorts of potential and actual causes are operating on organisms and populations thereof at any given time; some of these are more obvious than others. The DNA molecule, for example, represents a program operating toward the end--its final cause, one can truly say--of producing an organism fit to operate within a pre-designated ecological role, and capable of reproducing itself (note, moreover, that the final cause of DNA is not merely its reproduction to create more DNA: were this the case, then we would all still be viruses or their like). Consider, however, that this "program," as it now operates, has evolved in response to many impinging forces that manifest themselves not at the level of the molecule itself, but at different levels of organization such as entire organisms, populations, and ecosystems. Many of these forces impose absolute limits on form and function as well; DNA-based life forms can exist only within certain absolute limits of temperature, pressure, throughput of atomic or molecular substances, etc., etc. Further, and importantly, what is "absolute" at one point of development might not be so at a later time.
This last point has been but little considered either by philosophers or by scientists. There is in fact a whole world of difference between conditions "in which the events and structural results of a later time are viewed as to one degree or another preordained at an earlier time" (as stated above), and such conditions as might be specified at "some earlier time." In Aristotelian settings such as the famous example of the creation of a sculpture (the end cause of the sculptor's efforts), "an" earlier time is always specified: in that instance the point at which the sculptor conceives of and begins to plan the execution of his work. In the case of nonconscious entities, at any given point there may be changes in the environment that hasten the evolution of a particular new form; later, the descendants of that form may face changed conditions in the environment or biological community that prevent their further existence. Some of these causes might be easy to contextualize within simple survival of the fittest thinking, but others may be linked to deeper structural causes and necessitate the taking into account of more complex, environment-seated, inertias.
On several occasions (Smith 1984, 1986, 1989, 2004, 2005) I have discussed a "geographical" model of adaptive change in which systems concepts are used to deduce an evolutionary process more closely tied to community/resource development trends than to phylogenesis per se. Here, I should like to describe some additional ideas related to resource utilization that might be helpful to the investigation of complex processes. The main model developed here originates with a point touched on in one of the other works. In a note (Smith 1986, pages 272-273) I commented that certain "ubiquitous" environmental resources should not be considered "progressive" selection factors through the "two sector" (biotic and abiotic realms) model described there, as these presented only a condition for existence, and not for change. I will next elaborate on that point by developing some ideas designed to dynamize the present static interpretation of the relation between "niche" and "habitat."
3. The Hutchinsonian Niche and a Typology of Niche-Defining Relationships
In what has become a classic of the ecological literature, Hutchinson (1957) developed a philosophical context for the niche concept. In Hutchinson's words (p. 416), "an n-dimensional hypervolume is defined, every point in which corresponds to a state of the environment which would permit the species S1 to exist indefinitely. For any species S1, this hypervolume N1 will be called the fundamental niche of S1." Hutchinson proceeds to generalize these comments in such a way as to establish a model of the "ecological niche" (p. 416):
"...the fundamental niche may be regarded as a set of points in an abstract n-dimensional space. If the ordinary physical space B of a given biotop be considered, it will be apparent that any point p(N) in N can correspond to a number of points p(B) in B, at each one of which the conditions specified by p(N) are realised in B. Since the values of the environmental variables x1x2...xn are likely to vary continuously, any subset of points in a small elementary volume N is likely to correspond to a number of small elementary volumes scattered about in B. Any mean free paths of any animals under consideration is likely to contain points corresponding to points in various fundamental niches in N."
Through this approach, Hutchinson attempted to wed the facts of "commonness and rarity" (Preston 1948) in physical space to the competitive exclusion principle of Gauss and Volterra. The goal was apparently to shift our understanding of the concept "niche" toward a neo-Kantian one in which biological space was seen as the realization of competition among individuals and populations under particular locational conditions; earlier, the more typical approach had been to view the niche as a quasi-physical reality into which the organism "fit." Hutchinson's model has been a productive one, encouraging more interactive dialogs among workers in such varied fields of study as animal behavior, population biology, population genetics, and evolutionary and physiological ecology.
The adoption of the Hutchinsonian model of the niche has had a side-effect, however, that is less fortunate. In shifting their attentions to this new interpretation, ecologists inadvertently created a rift between population and frame of reference that extends beyond the simple differentiation between niche and habitat. When the concept of "niche" is expanded to anticipate all interactive aspects of an organism's existence, "habitat" is tacitly demoted to the static, measurable characteristics of geographic location. The problem with this separation is that along with it follows a view of space and the events that occur in/as space that oversimplifies the way these two things are related. From this view, how can it be distinguished with any confidence to what degree the population "defined" by a particular hypervolume associated with a given habitat and time is a product primarily of the properties of that hypervolume, or of that, or past, habitats? Hutchinson's approach necessitates parallel studies of two distinctly defined entities of which one, habitat, we can have (in Kantian terms) no direct knowledge because it is defined in a manner that excludes it from the system of interest. The Hutchinsonian understanding is incomplete to the extent that it identifies no explicit means whereby "niche" and "habitat" can be related in an interdependent (evolutionary) sense; i.e., in a mutually causal "external/internal factors" sense that may be instructive when we choose to focus on the possible long term implications of small scale, shorter term, processes.
I thus complain that while the hypervolume conceptualization may lead to quite useful insights within the context of community and evolutionary ecology studies, it offers less as a starting point for ecological and environmental evolution modeling. The Kantian framework can be quite effective when applied to events tacitly defined within a non-recursively evolving domain (note that Kant's philosophical system was in large part inspired by Newton's static, mechanistic interpretation of universal organization), being especially powerful in the context of cross-sectional analyses. This does not guarantee its appropriateness for longitudinal studies, however. How do habitats evolve? Do they represent simple reflections of the "sum of their parts," or do they exhibit their own characteristics of development that generate a distinct pattern of influence on the evolution and maintenance of niche relationships? To what kind of model do we turn for assistance in integrating location in space and time with the variously-scaled events occurring at that location?
In one of his lesser known papers, Hutchinson (1964, p. 931) points out the part that rates of availability of elemental resources might have on biochemical evolution:
"Evidently about a quarter of the (chemical) elements with 106-108 atoms per cell may have a (biochemical) function...although cobalt is enriched relative to nickel in liver, as in nearly all tissues of higher animals, over its concentration in the lithosphere, or for that matter in plants, it is still little more abundant than lead and less so than molybdenum. To use an element such as cobalt, the biochemistry of utilization must be reasonably specific. There are plenty of atoms of various kinds around in such concentrations that they could play a part as antimetabolites as well as significant functional roles in enzyme systems. It is possible that this sets the lower limits of concentration at which biochemically significant substances occur. There might be too many commoner accidental and potentially interfering materials around for any very important substance to work practically at 104 atoms or molecules per cell. The variety of elementary composition may thus set the standard of purity within which biochemical evolution has occurred."
Shortly thereafter, his discussion continues on to a consideration of limiting factors, but this is not placed within a framework which allows us to connect on equal terms those resource factors that are deemed limiting with niche evolution principles. We now introduce a typology that might be helpful in accomplishing such.
Hutchinson's level of attention in his 1964 paper is not clearly specified. At times he refers to the level of the individual, at others to population functions, and at still others to entire ecosystems. We shall look first at some matters concerning individual organisms. The individual, as a "negentropy machine" (Schrodinger 1945) continually in need of replentishing its sources of energy, may be viewed to respond to the characteristics of presence/absence of two fundamental kinds of resources: those that are ubiquitously distributed in time and space, and those that are not. (A "resource" may be understood here as any physical or biological factor/element paralleling Hutchinson's "environmental variable" concept.) We more commonly deal with the second kind of these, which may be defined here as a resource that can be obtained only through exertion/activity resulting from/in a directed movement in time and/or space. Thus, food (and all its component nutrients) is a non-ubiquitous resource for a terrestrial animal (because it must in one manner or another be actively sought), and water and temperature are non-ubiquitous resources with respect to a terrestrial plant (because a plant's physiology/morphology must change to adapt to temporal variation in availability of water and optimality of temperature).
By contrast, a ubiquitous resource may be understood as one whose processing is necessary to the persistence of the organism (even at rest), that is ubiquitous with respect to the organism's sphere of influence, and that need not be actively sought. There are probably no absolutely perfect examples of this class of resources, but rough examples include air for surface terrestrial creatures, water for aquatic ones, and gravity for both. In each case (and under all but extraordinary circumstances), the resource is available to an extent that demands no special strategies for obtainance.
In distinguishing between these two classes (or, if one prefers, "extremes") of resource type, four initial things have been accomplished. First, all resources possibly relevant to an organism's maintenance of well-being have been marked; added together, ubiquitous and non-ubiquitous resources represent all that an organism can conceivably be thought to respond to over the span of its existence.
Second, it theoretically becomes possible to reduce all functions/activities of an organism to an energy budget associated with: (1) a "ground state" (its maintenance of metabolic solvency as a function of its moment to moment processing of ubiquitous resources), and (2) the time spent on, and movements/efforts directed toward, the procurement of non-ubiquitous resources. (See more on this subject in Smith 1986.)
Third, a joint space has been established in which ubiquitous resources and non-ubiquitous resources co-exist in an ecological sense, duplicating Hutchinson's niche hypervolume concept. (Further, a complementary relativistic space internal to the organism can be conceived on the same basis. The condition "being alive" can be considered equilibrial to the extent that it defines a fundamental kind of ongoing "solution" to the set of all relevant impinging stresses. This "solution" may be dissected into its internal components on the basis of the degree to which particular metabolic/physiological processes maintain continuous operation--life--within the organism.)
Finally, the resource base of all organisms has been distinguished in such a fashion as to permit more flexible modeling of changing relationships between populations of organisms and environment over time. Our particular focus here will be on how a population might be "trapped" into responding to long-term changes in relationships between ubiquitous and non-ubiquitous resources. While our definition of non-ubiquitous resources leaves us able to recognize changing input/output relationships over time and space as organisms cope with varying rates of resource availability, the overall position also allows us to consider the possibility of longer-term evolutionary effects caused by changes in ubiquitous resources.
4. Maruyama's "Second Cybernetics"
Magorah Maruyama's "The Second Cybernetics: Deviation-Amplifying Mutual Causal Processes" (Maruyama 1963) was a milestone work in General Systems Theory that contrasted systems operations under what he termed "the first cybernetics" with a new understanding, "the second cybernetics" (p. 164): "The difference between the two types of systems is that the deviation-countering system has mutual negative feedbacks between the elements in it while the deviation-amplifying system has mutual positive feedbacks between the elements in it." Maruyama provided a further context for the two terms by associating the first with morphostasis and the second with morphogenesis.
Maruyama portrays evolution as a deviation-amplifying mutual causal system. He begins by noting that such systems appear to behave in a fashion running contrary to the second law of thermodynamics, and that this fact for some time embarrassed scientists before the emergence of the positive reinforcement concept (Prigogine 1961, von Bertalanffy 1968, Nicolis and Prigogine 1977). He then points out four more specific aspects of the evolutionary process that can be understood as deviation-amplifying: mutation/environment relations, interspecific competition, intraspecific competition, and the effects of inbreeding. This leads him to characterize evolution as "phylogenetic morphogenesis," and to an argument integrating ontogenesis into that schema.
It is in connection with the last discussion that Maruyama introduces a model of organismal growth that operates on the basis of a few rules that are self-enacting during the development process. He imagines a grid superimposed on a two-dimensional field within which an organism of like dimensionality may develop (in an ontogenetic sense). By introducing a set of rules of development and starting with four types of "cell tissue" at known locations within the grid, he points out that he is then capable of determining all aspects of the further development of those "tissues" via a line of reasoning set in the deviation-amplifying mode. This approach, he claims, has two advantages. First, it is a deterministic model, allowing for an inequivocal solution that is the same for all identical sets of initial conditions. Second, it is more efficient. The idea here is that when "the rules (of system evolution) are unknown, the amounts of information needed to discover the rules is much greater than the amount of information needed to describe the rules. This means that there is much more waste, in terms of the amount of information, in tracing the process backwards than in tracing it forward" (p. 174).
Despite the basic appeal of these ideas, they contain some philosophical restrictions making their translation into functional evolutionary models difficult. Perhaps most central is the problem that evolution involves more than a preset expansion process. Maruyama's simulation begins with a deterministic statement about the relative positions of the initial components of the system, but suggests no rationale for those locational choices (or the relation of the components to them, for that matter--a fundamental error that was instrumental in sinking the logical positivist school). Thus, the initial conditions of all developing subsystems must be identified and located a priori. In theory, this can be accomplished easily enough for an entirely deterministic ontogenetic system; rules could be included which would initiate new tissue types under the right sets of deterministic spatial configurations. This does not, however, clarify the evolutionary process which resulted in these new governing rules themselves because, using Maruyama's own reasoning, the amount of information needed to discover the evolutionary process behind them must still be greater than that needed to set them out. It seems preferable, therefore, to deal directly with the rules of evolution first, and then to deduce rules--of ontogenesis, for example--from these. Such was the trend earlier in the history of the development of evolutionary biology, but with time the goals of explanation have narrowed, and the pendulum has swung in the other direction.
5. Mutually Causal Environment-Organism Systems
Supposing that we are interested in modeling evolution as a deviation-amplifying mutual causal system--and I believe Maruyama is generally convincing in his argument that we should--what considerations should we emphasize as central in our rule-developing strategy? Evolution is to a recognizable degree a stochastic process involving the cumulative effects of the relationships among organisms and their environments. The general environmental framework of this process is well expressed in the following passage (Carson 1969, p. 76) pertaining to landform evolution:
"A system may achieve equilibrium between form and process (assuming that the external variables which control the processes do not change) almost immediately in some cases; in other instances, the system may proceed so slowly towards equilibrium that an evolutionary approach is necessary to understand the nature of the system at any one point in time. In the situations where a system rapidly achieves equilibrium between form and process, an evolutionary model is unnecessary and a complete understanding of the nature of the system is furnished by a knowledge of the way in which the equilibrium pattern depends upon the external variables. An exception occurs when the outside variables themselves change through time in a systematic manner: although it is still possible to understand the nature of the system at any one point in time by references to the current state of the controlling variables, a more complete explanation is afforded by setting the system in a historical framework."
For any given organism the sum of its world amounts to itself plus the environment external to itself. Over the span of its lifetime, both it and that environment change, albeit in ways and at rates that are peculiar to each. It will change as a function of growth, aging, and a variety of factors involved with the sum of its life activities. The environment will change as a function of the changes associated with the growth, aging, etc. of its components at any given slice of time, and with the cumulative results of the effect of lag of indirect causal forces spread among its components over time. We should be careful, it seems, not to confuse the two effects: that which differentiates organism from environment with respect to a particular organism is for any other organism simply a relationship contributing to the overall complexity of its environment. Organisms are distinct from their environments only in hindview, historically. If it is our desire to develop a general model of evolutionary process into which the idea of mutually causal processes involving organisms and environment is integrated, therefore, we must develop some way of distinguishing the two in a way that at once sees them as a joint space at any given point in time, but as separate components over the whole of time. We can be aided in this goal by the differentiation made earlier between ubiquitous and non-ubiquitous resources.
It should be apparent that any resource limited in space/time that can be processed in a manner favoring the preservation of living things will be competed for among those living things with the potential for adding it to their resource base. Thus, all non-ubiquitous resources are by definition competed-for resources. Further, all ubiquitous resources that are rare enough and that are fundamental to life processes will also be competed for. A reasonable example of the latter type is carbon dioxide, the relative rarity of which represents an immediate limiting factor to the overall rate of photosynthesis. This leaves one remaining class of resources about whose possible effects we need worry: those that are ubiquitous and present in such amounts as not to be competed for. Atmospheric oxygen provides a reasonably good example of this type. We cannot ignore this class of resources as a factor potentially influencing evolution, however, as it is necessary to consider the possibility that the presence of too much of such a substance might under certain conditions pose an unpredictable influence on physiological equilibrium. We might expect this to be especially likely in the circumstances of resources whose ambient levels increase slowly, but steadily, over long periods of time; i.e., such a trend might impose a constraint on a form's further evolutionary potential by creating an unsolvable contradiction between the goals of physiological/metabolic adaptation and ecological adaptation at some given point.
Admitting to the possibility of the above scenario, we can imagine an evolutionary model integrating probabilistic and deterministic elements which circumvents the criticism of the Maruyama model discussed earlier. It is generally agreed that adaptation is an opportunistic process that tends to lead in the long term sense toward increased specialization within individual lines. Specialization is largely a one-way street; i.e., adaptation occurs in response to the conditions of a given time and place, and once imprinted on the population's organizational theme represents a constraint on its range of possible future states (a theme well explored by Wiley and Brooks 1982). This should be true whether adaptation is occurring in response to competition for ubiquitous or non-ubiquitous resource types. In the latter case, the characteristics of the population are shaped as a function of competition for the heterogeneously-distributed resources of its surrounding environment. Should a given population become quite specialized as a result of long-term adaptation to a narrow range of community/environmental conditions, and should those conditions rapidly dissolve, the usual implication is extinction. This chain of events is easily dealt with within the Hutchinsonian framework, further useful dimensions being offered by the latter's connection to the well-developed concepts of natural selection, competition, etc. The constraints implied by adaptation to ubiquitous resource processing, however, have not been explored to the degree their probable importance merits. (Part of the reason for such avoidance is undoubtedly the fear of falling into either teleological or overly deterministic modes of thinking, as mentioned earlier.)
Since a population's equilibrium depends on operating within a given sum of environmental/ecological conditions, both ubiquitous and non-ubiquitous resources must somehow be involved in shaping the adaptations of its elements in an evolutionary sense. I would argue, however (as noted earlier), that while it may be a condition of existence for an organism to be able to process each kind of resource, this fact alone does not mean that ubiquitous resources, at least, will necessarily have any significant influence on the rates at which new characters are retained; this, in view of the more immediately impinging environmental/ecological stresses. When the rate at which qualitative/quantitative adjustments take place in ubiquitous resources involves a much longer time scale, the direction of specialization must be largely a function of non-ubiquitous influences rather than ubiquitous ones (i.e., the classic Darwinian fitness model).
Regardless, all novel physiological strategies initially evolve under some given levels of ubiquitous resources, and as the latter change over geological time, so too must the organism (i.e., population) if it expects to continue to compete. Along these lines, one can imagine a scenario involving such change that might lead to a hitherto unexplored influence on the process of extinction. Specifically, a physiological plan appropriate to given conditions of ubiquitous and non-ubiquitous constraint at time t might cease to be viable when it is no longer able to sustain the organism as some critical level of a slowly changing ubiquitous resource is encountered at time t + n. The mechanism may be summarized as follows.
Hutchinson (1964) touches on the matter of physiological constraints on organic change when he refers to the lowest concentrations of elements to which productive reaction can take place. I believe it may be of yet more interest in the sense of long term function to consider the possible effect that certain elements reaching highest concentration allowable might have on an organism (more properly, a population). Although organisms are reasonably well protected from the negative effects of unwanted chemical reactions by their physiological specificities, it seems that under certain conditions such protection may become difficult to maintain. It is well known that even generally beneficial elements or compounds may sometimes produce negative side effects when rates of intake or exposure become too high. While one would expect most serious problems of this type to be alleviated over time through selection, this assumes that it is possible for the organism to do so without compromising already necessary structure. If conserving that already necessary structure depends on maintaining--or even increasing--a level of utilization of the element/compound that is generating the negative side effect to begin with, selection may not be able to overcome the problem (or if it does, only temporarily).
The possible relevance of the evolution of oxygen levels in the atmosphere to the present discussion is particularly interesting to consider. It is difficult to deny that the long-term historical increases in atmospheric oxygen levels have made possible energy-exploiting diversification trends that otherwise likely would not have taken place (see, for example, Dudley 2000, Berner et al. 2003). But the biochemistry of oxygen has two distinct faces: while fundamental to the ultimate purpose of respiration, oxygen also produces interactions having any number of potentially detrimental effects on cellular operation that must, at the same time, be guarded against (Caughey 1979, Lane 2002, Vallyathan et al. 2004). Importantly, there is no reason to suppose that the magnitude of all such effects should be invariant with respect to the changing constitution of the atmosphere; i.e., what might not be "too much oxygen" for an organism at one stage in geologic time very well might be at a later stage. Thus, we may end up with a selection regime that is working at cross-purposes: it will not suffice, one supposes, to simply adapt to the problem by reducing efficiency of oxygen utilization in the relative sense, as this is bound to reduce the organism's ecological efficiency as well. In many instances, extinction might be regarded (in retrospect, at least) as being preordained to the extent that a group's initial direction of (physiological) adaptation would later prohibit it from maintaining a productive selection regime, given the levels of interference with fundamental metabolic process brought on by an unpredictably rising ubiquitous influence. Extinction under these conditions would be, in a real sense, subservient to a final cause connected with the increasing levels of the resource.
This model of interacting constraints potentially permits us some new flexibility in understanding the processes of speciation and extinction. While we might continue to consider the process of adaptation--of whatever type--entirely opportunistic (stochastic) in its earlier stages of unfolding, we can begin to regard later adaptive programs and extinction as rather more complex kinds of events. Extinction might still be linked to a population's inability to modify appropriately in response to changes in its ubiquitous and nonubiquitous resource base, but the reason for this inability, I suggest, might usefully be dissected into its "necessary" and "sufficient" components. The former will be associated predominantly with historical factors peculiar to the gene pool of the population; the latter, with environmental change.
Of special interest here is the fact that this model suggests the likelihood that extinction episodes will often be probabilistically hierarchical. There will be no way for an ancestral population to "know" beforehand whether a particular physiological strategy that works under then-present conditions will become ineffective under some future conditions. If the future stress devolves as a by-product of the necessary utilization of some ubiquitous resource, all related forms sharing the physiological strategy in question will be simultaneously affected, independent of ecological circumstances. If the strategy is only a slight variation on a generally more applicable theme, extinction may remove only one or a few closely related forms. If, on the other hand, the strategy is of considerable generality and sustained a major radiation, the extinction of a whole line of forms could take place--and quite rapidly, at least in geologic terms. This interpretation might even be extended to multiple lines, if their initial evolution was a function of analogous exploitations of environmental opportunities, or if important food chain or other relations were involved.
6. A Case Study Scenario: The Extinction of the Dinosaurs
We might next begin to look for specific examples of such conditions, and to consider evidence bearing on related interpretations. This is not a matter to be explored adequately in one short paper, but we may reasonably consider at least one fairly obvious, and famous, application.
It is now widely accepted that levels of atmospheric oxygen have fluctuated considerably during Phanerozoic times (Brancazio and Cameron 1964, Cloud 1968, 1976, Berner and Canfield 1989, Berner et al. 2000, Graham et al. 2000, Lane 2002), leading to the present level of approximately twenty-one percent of the atmosphere. Historically, four general regimes of development can be recognized during Phanerozoic time: (1) an early to mid Paleozoic period at around, or less than, fifteen percent (2) a later Paleozoic spike with values approaching thirty-five percent, with a subsequent drop across the Permian-Triassic boundary to early and mid-Triassic levels of less than fifteen percent (3) a general rise from mid-Triassic times to levels of about twenty-five percent across the K-T boundary (4) a subsequent gentle decrease to present levels.
Years ago Bakker (1971, 1972) and Ostrom (1969) independently arrived at the conclusion that dinosaurs might have been warm-blooded creatures. Bakker, Ostrom, and the many later proponents of this view have been able to back it with some fairly convincing arguments; nevertheless, it seems that the ensuing discussion has become mired in an increasing number of sub-explorations for which multiple lines of logic seem to hold, or that are untestable, or both. Among these are those concerned with dinosaurian extinction. Literature on the extinction of the dinosaurs is now very extensive, and no review will be attempted here (see Powell 1998, Ellis 2004, Fastovsky et al. 2005, as a start). Suffice it to say that the matter is still not fully resolved, the sensational effect of the "cosmic calamity" asteroid collision model (Alvarez et al. 1980) on our thinking notwithstanding. At present, discussion has progressed from a general acceptance that the collision event did in fact take place to various considerations of how long, and for what reasons, the dinosaurs (and other groups) might have lingered on afterwards.
Prior to the introduction of the collision model, much of the endothermy-related discussion on the K-T mass extinction event focused on the problem of how a warm-blooded dinosaur might have been able to maintain a minimum level of metabolic activity under changing environmental conditions (especially those related to cooling trends typifying the mid and late Cretaceous). This matter has now largely taken a back seat, but seemingly equal attention might be paid to a different question: given steadily increasing atmospheric oxygen levels over this period, might the later Cretaceous dinosaurs have experienced more and more difficulty keeping within maximum allowable metabolic tolerances, and begun to adopt behaviors, evolve structures, and change range accordingly?
For the sake of discussion let us suppose that, given relatively low levels of atmospheric oxygen and the general advantages associated with evolving endothermy, two means of striking a flexible balance between heat loss and retention are effective: (1) small size (to promote heat loss under high temperature conditions) combined with external insulation (to promote heat retention under low temperature conditions), and (2) large size (to promote heat retention) combined with internal or negligible insulation (to promote heat loss). Let us further suppose that the second strategy, while workable under low ambient oxygen level conditions, has no guarantee of working well--or at all--under much higher level conditions. If creatures of the second class were overspecialized to the point of being unable to simultaneously decrease in size, evolve an external form of insulation, and maintain a certain range of body temperatures, an irreconcilable stress might be exerted. In sum, I entertain the idea that the dinosaurs, though effectively specialized to capitalize on the environmental conditions of an earlier time, may have lost out in the long run when one element of those conditions (perhaps, specifically, oxygen levels) modified itself out of their later range of potential adaptability. This brand of "final causes" model differs from traditional treatments regarding adaptability in its assumption that some circumstances of macro-organization are based on a more adaptable kind of resistance than others to certain negative influences on micro-level function; i.e., especially as such influences increase in relation to increases in a resource that contributes fundamentally to system organization.
A number of lines of evidence actually support the plausibility of such a scenario, particularly with respect to the oxygen levels change interpretation:
(1) The rate of dinosaurian disappearance was quite rapid but some (Christensen and Birkelund 1979, McLean 1978, Russell 1982) initially argued not entirely abrupt; subsequent clarifications involving stratigraphic unit clarification, radiometric dating, fossil assemblage censusing, statistical sampling difficulties, etc., moreover, have remained inconclusive on this point (Archibald 1996, MacLeod 1996). Unless the extinction was virtually instantaneous, even a short period of demise of some thousands of years is probably enough to support the kind of interaction proposed--especially considering the other major environmental changes that took place during this period, the rather narrow range of body temperatures within which even a major taxon of endotherms operates efficiently, the probable "domino effect" that would likely occur in the food chain once some critical number of species became involved, and the possibility that the upper tolerable limit of stress on the organismal system may have been a fairly acute tipping point by then easy to permanently transgress.
(2) Almost all of the dinosaurs were at least relatively large, some indeed being giants. All, therefore, would have been affected to one degree or another by the size/physiology trade-off problem envisioned.
(3) If birds are derived from dinosaurian stock, as is now largely accepted (Bakker and Galton 1974, Paul 2002, Currie 2004), the fact that they survived and their larger cousins did not can be attributed to their early adoption of smaller size and feathers as physiological aids to exploiting an aerial niche. Body temperatures of birds in general are higher than those of mammals, and might in part be linked to their affinity with the reptilian version of endothermy and its evolutionary limitations within a nonequilibrial atmosphere.
(4) The subordination of mammals to reptiles throughout the Mesozoic might be attributed in large part to this posited link between oxygen levels and the relative efficiencies of the groups with respect to their heat retention/loss trade-off strategies. Once reptiles gained control of the food chain, it may have been difficult for early mammals to specialize to an extent supporting size increase, even when this presumably became possible later in the Mesozoic. This is not to say that there might not have been many additional contributing factors, of course: perhaps, for example, early mammals lacked tail musculature appropriate for holding up a tail large enough to provide the counter-balance needed to offset increased body weight, as was suggested years ago by Dodson (1974).
(5) The earliest known dinosaur fossils are of Carnian age, late Triassic (Benton 1997). Even if this underestimates the group's actual origination period, this would seem very unlikely to be before the middle Triassic. At this point atmospheric oxygen levels were just beginning to recover from their lowest values, some thirteen percent of the atmosphere, following the end of the Paleozoic. Apart from a temporary dip at the Triassic-Jurassic boundary (coinciding with another mass extinction), therefore, and judging from the figure reproduced in Lane (2002, p. 83), the entire period of dinosaurian existence coincided with a persistently rising atmospheric oxygen content, to a peak of over twenty-five percent at the K-T boundary (a total increase of about one hundred percent over original values):
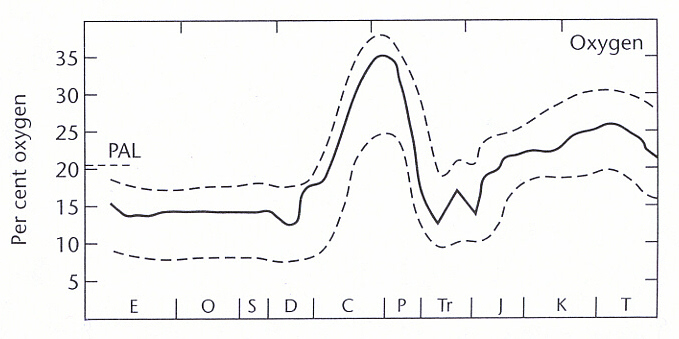
(6) Interpretation of the continuation of the cold-blooded reptiles becomes relatively straightforward, as they were progressing according to an entirely different physiological game plan.
(7) The continuing gigantism exhibited by dinosaurs might thus be interpreted as the simplest response to lack of external insulation, with the evolution of great size as a means of balancing heat loss and retention. The world cooling of temperatures in the Late Cretaceous might be viewed as one of those ecological stresses to which the dinosaurs had to maintain a viable adaptive response, and under the conditions that response would likely have been, for reasons of historical inertia, maintenance of large size.
(8) It is interesting and perhaps significant in this context that the ocean-going mammals of today, whose insulation is primarily internal and whose average size approximates that of the dinosaurs, are largely (though admittedly not entirely) relegated to cool or cold water environments. Note that the present existence of large mammals does not in itself contradict the model of dinosaurian extinction explored here. Rather, the interpretation is that early mammals, in maintaining small sizes, retained a less specialized state that allowed them to later evolve a physiological flexibility of large size under higher-oxygen conditions. This does not mean, however, that in general very large size has not become an increasingly less viable physiological adaptive strategy to ecological conditions; one perhaps, that necessitates counter-strategies such as association with cold and/or aquatic habitats. On land, maximum size among the mammals was apparently reached quite some time ago (in the Oligocene, with Paraceratherium)--during a period of gradually descending oxygen levels. Moreover, of the three groups of largest present day terrestrial mammals (elephants, hippopotami, and rhinoceroses), two habitually utilize water as a means of cooling off, and all three are in the late stages of a long term decline in overall diversity/ecological significance.
An important related question is whether there might have been a trend among Cretaceous dinosaurs toward occupation of more and more temperate and/or aquatic habitats. The finding (and subsequent confirmations) that non-migratory hadrosaurs and other dinosaurs existed on the North Slope of Alaska circa 76 to 66 million years ago (Brouwers et al. 1987) certainly represents positive evidence on this score, and it would be interesting to see a review of all relevant data (my impression is that there seem to be fewer tropical zones dinosaur fossil records for late Cretaceous times than from earlier periods). And, although I am unaware of any data bearing directly on changing aquatic habits of dinosaurs, the well-known study by Archibald and Bryant (1990) indicated that of all vertebrates living in eastern Montana at the time of the K-T boundary, up to ninety percent of those forms that lived in aquatic environments survived into the Cenozoic, whereas the parallel figures for terrestrial forms were as low as ten percent.
(9) Although it is still not impossible that a statistical artefact involved in the sampling of paleo-populations is the cause, it would appear (Archibald 1996, MacLeod and Keller 1996) that dinosaur diversity (as well as that of some other groups) had already peaked some three million years earlier and was in considerable decline at the time of the K-T boundary, suggesting an extinction process involving factors beyond those connected with the collision hypothesis alone.
(10) Some of the morphological peculiarities of the dinosaurs might be interpreted within this context. The evolution of "crests" (e.g., Parasaurolophus and Corythosaurus) and "sails" (e.g., Spinosaurus), etc., might have represented strategies, at least in part, for reducing the stress associated with temperature regulation under conditions of large size. (This interpretation has existed for some time, but to my knowledge has not been related to other than more immediate causal conditions, or to the old concept of "evolutionary senescence"--which itself takes on some new dimensions through the model espoused here).
(11) It should generally be observed that the wholesale extinctions of some families and orders seems to suggest a priori some causal agency connected to weaknesses common to the basic morphological/physiological plan of whole groups of organisms more than it does a tie to specific environmental events (a "sufficient" as opposed to "necessary" causes kind of interpretation).
(12) It is interesting and quite possibly not coincidental that the distribution ranges of many of the more primitive living representatives of ancient groups seem to coincide with relatively oxygen-poor environments. This is particularly true with respect to mountainous areas. Among the terrestrial vertebrate examples that can be cited are aplodontid rodents (northwestern United States), ascaphid frogs (northwestern United States), ichthyophiid caecilians (southern India), tropidophiid boas (Middle America), uropeltid snakes (southern India), Zaglossus, a tachyglossid (New Guinea), Micropotamogale, a tenrec (West Africa), Southern Appalachian salamanders (Plethodon and others), and caenolestid marsupials (Andean South America). Adding other groups and habitats (e.g., swamp and subterranean environments) would produce quite a long list. Other kinds of constraints permitting (e.g., stability of environment), one would expect this kind of association to develop on the basis of present thoughts, as such habitats would sponsor atmospheric oxygen levels more closely resembling those of earlier periods. In this instance regression models relating relevant current environmental variables to geological age of ancestral form can be imagined that might shed light on the posed relationships.
For the moment I advance the preceding points as food for thought only. Certainly there are many kinds of biotic patterns studies that can be imagined that would shed further light on the subject. The "cosmic calamity" interpretation itself might benefit from related efforts if it could be shown how such an event might have temporarily increased oxygen levels or set other relevant chains of effect into operation.
Speaking of other relevant chains of effect, it should finally be noted in the context of this particular discussion that longitudinal scenarios involving oxygen and posing other than simple heat balance problems can--and should--also be entertained. As pointed out earlier, oxygen is potentially a physiological poison presenting dangers from which organisms must guard themselves. The significance of oxygen to bodily function is a very large subject, and a good many other possible causal scenarios might be imagined that could be relevant in this particular context. It seems difficult to directly connect Late Cretaceous mass extinctions of plants to the specific mechanism I entertain above, for example, but this doesn't necessarily mean that other conditions of oxygen processing might not be related to their, or animals', demise. Suppose, for example, that rate of ATP-ADP conversion is strongly linked to the degree to which an organism can protect itself from overutilization of oxygen. Or, as another scenario, suppose that the same is true with respect to generally increasing levels of toxic oxygen compounds such as ozone. Either of these mechanisms also fits well into the general model of lagged interactions described here. These may not constitute "explanations" at this point, but they are nevertheless not improbable a priori.
7. A Simulation Model
The notion that a general class of "lagged effect" causalities such as described above may exist and affect the evolutionary progression may be a difficult one to test outright given current levels of knowledge, but in addition to the reasoning out of suggestive relationships one might turn to simulation studies as a means of investigation. In recent years considerable effort has been devoted to developing models simulating the conditions under which earth's atmosphere might have evolved (e.g., Payton and Arrigo 2000, Berner et al. 2003, Hansen and Wallmann 2003, Bergman et al. 2004), but the approach advocated here differs considerably in its desire to do more than just set rules on the basis of positive and negative feedbacks (per criticism of the Maruyama simulation stated earlier), but to examine how a "final cause"--in this case a ubiquitous resource whose own changing levels may block future potentials in unpredictable ways--might affect rates of phylogenesis and extinction.
Much more involved and realistic sets of simulation rules might be developed than those I am about to describe, but there is no need to get bogged down in details to make the main point. A simulation field is imagined with a finite but unbounded grid surface of continuously interfacing cells (each in the shape of an equilateral triangle) such as might cover the outside surface of a dodecahedron. Upon this field unfolds a constrained contagious diffusion process meant to simulate the relationship of phylogenesis to environmental (more specifically, non-competed-for ubiquitous resources) evolution as a progression of interdependent steps of change. The rules of the simulation call for the step-by-step filling of the field by elements of two distinct classes, Y and X. Elements of class Y are "organic units"--taxa--whose "phylogenesis" is represented as a branching, contagious diffusion process away from an arbitrarily chosen starting point within the field. Elements of class X are "environmental units" whose more passive, derivative, evolutionary development is expressed in a simple random process of addition. The "non-X" portion of the field represents a "potential niche space" within which, when filled (and at any given point of its filling), may co-exist some finite (but, imaginably, very large) number of organic units. The primary organizational constraint on system evolution is that every time an element of class Y is added to the field deterministically (through contiguous diffusion), an element of class X is added at random. When the random placement of a new class X element involves a cell already occupied by a Y element, the latter is replaced with the former, an "extinction" event being recorded. A simulation field of relatively few cells eventually fills up, such that there are no vacant contiguous cells that class Y elements may "evolve" into, and the simulation ends.
The purpose of the simulation is to provide a rough analog to the general set of events that will emerge as a dynamic, opportunistic organic system evolves in response to the constraints imposed on it by its own history and by an environment: (1) that is stressful and limiting in an immediate sense, and (2) that will change unpredictably in the future. Thus, certain directions/strategies of diffusion (evolution/adaptation) are blocked by the existence of ambient factors that make these impossible initially; moreover, in some cases even those that are possible at one time end up being not so under later conditions of change in the environment (an "extinction" resulting).
Thirty simulations of this kind were executed, and some results tabulated. The number of steps required to achieve completion varied considerably (12 to 31), but most of this variation resulted from minor late-stage adjustments, and the simulations otherwise proceeded in reasonably parallel fashion. In Figure 1 I have plotted the mean number of each main kind of event (the addition of elements of each class and number of extinctions occurring) at each step of the simulation. Of greatest interest is the emergence of a decided lag between the maximum number of Y or X elements added in a step and the maximum number of extinctions occurring as a result of addition of X elements. This illustrates, albeit simply, the way in which an evolving system may be affected by the cumulative results of earlier-occurring events.
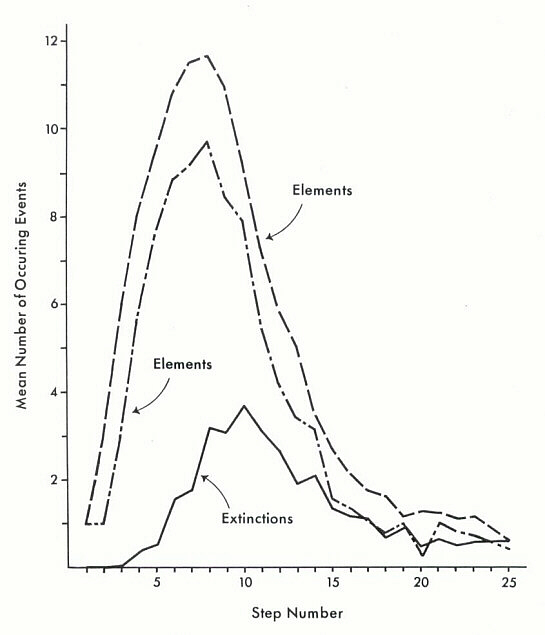
Figure 1. The mean number of elements Y and X added, and "extinctions" occurring, during each step of the simulation (based on thirty runs). See text for discussion.
In Figure 2 trend lines connect the total number (through twenty simulations, in this case) of Y elements (from each step of addition) remaining after each following step. A variety of rates of extinction (trend line slopes) emerge, depending on the original stage of introduction of organic unit. We find, for example, that despite the fact there were on the average far more elements added during step seven than during step ten, more step ten-added elements usually remained by the end of the simulation. Another trend that was quite apparent through these thirty runs was the tendency for a step involving a relatively large number of "extinctions" to be followed by/preceded by one involving a relatively low number (as assessed by a modified number-of-runs test on standardized data).
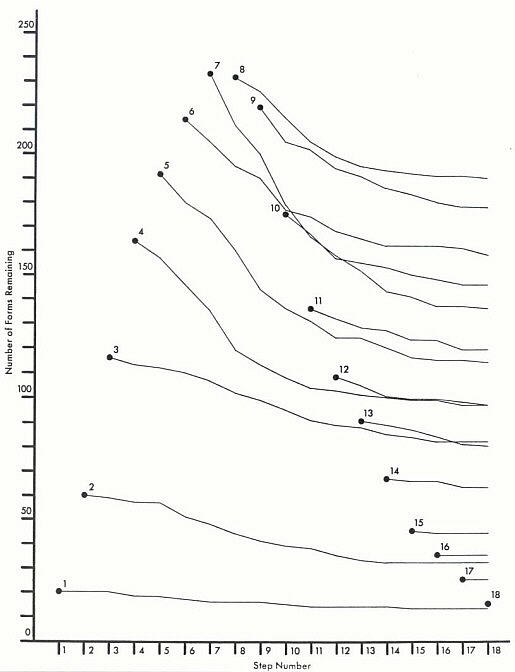
Figure 2. Total number of Y elements remaining after each successive step of the simulation (based on twenty runs only). See text for discussion.
Obviously, more complex and realistic simulations of this sort could be designed through adjustments to the specified implications of an extinction event. For example, one could set things up such that during a replacement episode all Y elements derived from a particular extincted Y element are also eliminated from the field (a "mass extinction"). Or one could stipulate a probabilistic function of removal of descendants such that less remotely derived forms are more likely to be removed. Or the converse. One could also attempt to run the diffusion process one extension at a time rather than as I have done (in discrete, exhaustive steps). It would be interesting to see whether any reasonably simple combination of assumptions can lead to the generation of trends approximating the historical pattern of real world extinction rates. (More involved simulations integrating geographical factors/constraints could even be designed that would permit regional differentiation modeling, and thus biogeographical study.)
8. Two Further Applications
Lest the impression be gained from the preceding discussion that the points made here are intended for application to long-term earth history events only, brief mention should be made of two other contexts that invite a final causes kind of application.
(1) In the early 1980s I spent some time investigating the spatial trends associated with wildlife's ten-year cycle in northern North America (Smith 1981, 1983). I quickly became intrigued with the biogeography of the event--specifically, how whole species populations could undergo synchronized, periodic cycles over time and space. For the past twenty-five years or so the main study approach to the problem has been through demographic analysis of food-chain relationships among the species populations involved (Berryman 2002), but these efforts have really brought us no closer to understanding how the whole phenomenon got started. I suggest that a solution will be obtained only when the larger scale relationships are better understood. Through my early literature review I repeatedly encountered the observation that at times of high population snowshoe hares were often afflicted with a disorder of unknown etiology that affected the liver; it appeared to be stress-related, and has frequently been referred to as "shock disease." Considering this, and the fact that the entire northern zone where the cycles take place is exactly situated where the terrestrial surface interface is least protected from extraterrestrial radiation bombardment by the geomagnetic field, I began to wonder whether "shock disease" might most fundamentally be caused by a photosensitivity-related disorder, and in turn how the Pleistocene biogeography of the region might be related to cycle generation. Perhaps as snowshoe hare populations were pushed southward several times during the Pleistocene ice extensions, they lost both their resistance to the photosensitive pigments once ingested in their diet, and their tendency to develop a white winter coat (neither stress any longer being a selection factor at more southerly latitudes). Returning northward during interglacials, a selection regime would follow favoring the development of light-colored individuals for concealment reasons; it is known, however, that photosensitivity-related disorders such as eczema (which duplicates some of the "shock disease" symptoms, for example skin lesions) particularly affects animals with light-colored skins and hides. Perhaps, as has been suggested by Sinclair et al. (2003), two population morphs have developed in conjunction with this set of contradicting pressures; in theory, until the gene pool identifies a solution to both selection pressures, we may expect continuing cycling (the details of which can then be understood through the demographics of life cycle and food chain relations). Note that here again we have a possible instance of a "ubiquitous" factor (irradiation) generating a condition through the imposition of intervening circumstances.
I have hung onto this interpretation for over twenty years, but until now have seen no way to make it public. Still, some other researchers, at last, seem to be moving in directions not far removed from this model. Many studies have attempted to link quality of food resources to the phenomenon, and at least one (Selas 2006) has gone so far as to suggest a relationship between sunspot activity cycles and a UV-B-induced plant stress on the population. I would suggest that researchers give some attention to the distribution of a possible photosensitizing agent as related to the characteristics of the strength of the geomagnetic field at high latitudes; among other relationships, it is stronger over the Canadian arctic than over Siberia, a possible reason why hare cycles are less pronounced in the latter area despite the apparent ideal ecological conditions and population webs.
(2) My most extensively considered example of how a final causes kind of framework might influence evolutionary processes is my earlier-mentioned work on the forces underlying biogeographical range change (Smith 1984, 1986, 1989, 2004, 2005). In these analyses I have argued that natural selection represents the negative feedback element (the "deviation-countering" factor, in Maruyama's parlance) in evolution; operating on pre-existing variation it simply maintains system equilibrium by removing inefficient forms of variation. The positive feedback portion of the cycle operates as the adaptive suites of all populations engage themselves, in ever-changing ways (through locational changes and ecological involvements), with their environment; some of these engagements eventually lead to short-lived trends of speciation, whereas others promote longer-lasting ones. On the whole, the longer-lasting ones have predominated, which suggests that the environment itself is organized in such a fashion as to channel the sum of these engagements toward generally higher levels of order. In the works just mentioned I have suggested that the spatially varying levels of availability of a very few fundamental resources such as water create a driving force in this respect, as populations will find it easier, even just by chance, to extend in geographical directions of their more optimal delivery because it will be easier to genetically select adaptive mechanisms to support engagement in these than in places where their delivery is more erratic. Populations of living things may be able to adapt faster than the changes that take place in the physical world, but this does not guarantee evolution in the broader sense. Thus, the environment becomes a kind of final cause--not to the degree that it forces particular adaptive structures (as was thought through the old environmental determinism models), but to the extent that it encourages unlimited refinements of biogeochemical cycling that generally support phylogenesis.
Again, it would seem that a number of other workers are moving slowly in this direction, though they continue to take a limiting factors-based ecological, rather than a biogeochemical cycles-based evolutionary, view of the resulting patterns of biodiversity. This may be seen especially clearly in the attempts to link solar energy availability, through various surrogates, to species richness and other elements of biodiversity (see, for example, Hawkins & Porter 2003; Hawkins et al. 2003; Kerr & Currie 1999; Kerr & Packer 1997). Some of these (e.g., Bhattarai et al 2004; O'Brien 1998; Currie 1991) have even specifically used potential evapotranspiration measures as their key indicators in such studies (as I did in 1984). These efforts, however, while methodologically sophisticated, have not attacked the main problem of how to link such indicators to an evolutionary framework, as was accomplished in the model developed in Smith (1984).
9. Final Remarks
While it is true that teleology and final causation are usually viewed as antithetical to scientific thinking, it may well turn out that complete appreciations of complex processes will never be possible until researchers stop relying on efficient causes models alone as their means of making headway. The degree to which evolution is in a multifaceted sense recursive--that is to say, both self-reinforcing and self-modifying--is still very poorly understood. The Hutchinsonian model of the niche, while potentially dynamic with respect to the operation, or even evolution, of organisms within a hyperspace, nonetheless fails to integrate actor and stage in such a manner as to facilitate explicit understanding of how each contributes to the evolution of the other. Evolutionary models such as Maruyama's "deviation-amplifying mutually causal" morphogenesis (Maruyama 1963) and Wiley and Brooks' "nonequilibrium thermodynamics" (Wiley and Brooks 1982) fair no better in this respect. True, these accept that there are external influences on phylogenesis (or even must assume such), but they remain incomplete in their treatment because no particulars regarding the possible lag effects of those influences is integrated into the understanding of the limitations under which biological systems function.
Through the "ubiquitous/non-ubiquitous" resources model discussed here it is possible to envision a biological history that derives from more than the naive assumption of a selection process dictated by immediate environmental opportunities and constraints only. Rather, the possible influence of joint conditions of internal and external control are acknowledged, the result being a kind of "push-pull" appreciation of causal influence. This writer is not the only person defending such kinds of thinking--proponents of the anthropic principle and gaia hypothesis (Barrow and Tipler 1986, Lovelock 1988, Margulis 1998) come immediately to mind in this respect, though their approaches are somewhat different. Whether we entertain full-blown teleology or something more subtle, however, I suggest that we will increasingly discover that there are forces operating on the biology-environment interface that are not understandable in efficient causation terms alone.
References Cited
Alvarez, L. W., W. Alvarez, F. Asaro and H. V. Michel. 1980. Extraterrestrial cause for the Cretaceous-Tertiary extinction. Science 208: 1095-1108.
Archibald, J. D. 1996. Testing extinction theories at the Cretaceous-Tertiary boundary using the vertebrate fossil record. In N. MacLeod and G. Keller (eds.), Cretaceous-Tertiary Mass Extinctions: Biotic and Environmental Changes. W. W. Norton, New York, pp. 373-397.
Archibald, J. D. and L. J. Bryant. 1990. Differential Cretaceous/Tertiary extinctions of nonmarine vertebrates: Evidence from northeastern Montana. In V. L. Sharpton and P. D. Ward (eds.), Global Catastrophes in Earth History: An Interdisciplinary Conference on Impacts, Volcanism, and Mass Mortality. Geological Society of America Special Paper 247, pp. 549-562.
Bakker, R. T. 1971. Dinosaur phylogeny and the origin of mammals. Evolution 25: 636-658.
Bakker, R. T. 1972. Anatomical and ecological evidence of endothermy in dinosaurs. Nature 238: 81-85.
Bakker, R. T. and P. M. Galton. 1974. Dinosaur monophyly and a new class of vertebrates. Nature 248: 168-172.
Barrow, J. D. and F. J. Tipler. 1986. The Anthropic Cosmological Principle. Oxford University Press, Oxford.
Benton, M. J. 1997. Origin and early evolution of dinosaurs. In J. O. Farlow and M. K. Brett-Surman (eds.), The Complete Dinosaur. Indiana University Press , Bloomington, pp. 204-215.
Bergman, N. M., T. M. Lenton and A. J. Watson. 2004. COPSE: A new model of biogeochemical cycling over Phanerozoic time. American J. Science 304: 397-437.
Berner, R. A. and D. E. Canfield. 1989. A new model for atmospheric oxygen over Phanerozoic time. American J. Science 289: 333-361.
Berner, R. A., D. J. Beerling, R. Dudley, J. M. Robinson and R. A. Wildman Jr. 2003. Phanerozoic atmospheric oxygen. Annual Review of Earth and Planetary Sciences 31: 105-134.
Berner, R. A., S. T. Petsch, J. A. Lake, D. J. Beerling, B. N. Popp, R. S. Lane, E. A. Laws, M. B. Westley, N. Cassar, F. I. Woodward and W. P. Quick. 2000. Isotope fractionation and atmospheric oxygen: Implications for Phanerozoic O2 evolution. Science 287: 1630-1633.
Berryman, A. A. (ed.). 2002. Population Cycles: The Case for Trophic Interactions. Oxford University Press, Oxford.
Bertalanffy, L. von. 1968. General Systems Theory. George Braziller, New York.
Bhattarai, K. R., O. R. Vetaas and J. A. Grytnes. 2004. Fern species richnesw along a central Himalayan elevational gradient, Nepal. Journal of Biogeography 31: 389-400.
Brancazio, P. J. and A. G. W. Cameron (eds.). 1964. The Origin and Evolution of Atmospheres and Oceans. Wiley, New York.
Brouwers, E. M., W. A. Clemens, R. A. Spicer, T. A. Ager, L. D. Carter and W. V. Sliter. 1987. Dinosaurs on the North Slope, Alaska: high latitude, Latest Cretaceous environments. Science 237: 1608-1610.
Carson, M. A. 1969. Models of hillslope development under mass failure. Geographical Analysis 1: 76-100.
Caughey, W. S. (ed.). 1979. Biochemical and Clinical Aspects of Oxygen. Academic Press, New York.
Christensen, W. K., and T. Birkelund (eds.). 1979. Cretaceous/Tertiary Boundary Events. Vol. 2. Proceedings. University of Copenhagen Institute for Historical Geology and Paleontology, Copenhagen.
Cloud, P. 1968. Atmospheric and hydrospheric evolution on the primitive earth. Science 160: 729-736.
Cloud, P. 1976. Beginnings of biospheric evolution and their biogeochemical consequences. Paleobiology 2: 351-387.
Currie, D. J. 1981. Energy and large-scale patterns of animal- and plant-species richness. American Naturalist 137: 27-49.
Currie, P. J. 2004. Feathered Dragons: Studies on the Transition from Dinosaurs to Birds. Indiana University Press, Bloomington.
Dodson, P. 1974. Dinosaurs as dinosaurs. Evolution 28: 494-497.
Dudley, R. 2000. The evolutionary physiology of animal flight: Paleobiological and present perspectives. Annual Review of Physiology 62: 135-155.
Ellis, R. 2004. No Turning Back: The Life and Death of Animal Species. HarperCollins, New York.
Fastovsky, D. E., D. B. Weishampel and J. Sibbick. 2005. The Evolution and Extinction of the Dinosaurs. 2nd Ed. Cambridge University Press, Cambridge.
Hansen, K. W. and K. Wallmann. 2003. Cretaceous and Cenozoic evolution of seawater composition, atmospheric O2 and CO2: A model perspective. American J. Science 303: 94-148.
Hawkins, B. A. and E. E. Porter. 2003. Relative influences of current and historical factors on mammal and bird diversity patterns in deglaciated North America. Global Ecology and Biogeography 12: 475-481.
Hawkins, B. A., E. E. Porter, and J. A. F. Diniz. 2003. Productivity and history as predictors of the latitudinal diversity gradient of terrestrial birds. Ecology 84: 1608-1623.
Hutchinson, G. E. 1957. Concluding remarks. Cold Spring Harbor Symposia on Quantitative Biology 22: 415-427.
Hutchinson, G. E. 1964. The influence of the environment. Proceedings of the National Academy of Sciences of the U. S. A. 51: 930-934.
Kerr, J. T. and D. J. Currie. 1999 The relative importance of evolutionary and environmental controls on boad-scale patterns of species richness in North America. Ecoscience 6: 329-337.
Kerr, J. T. and L. Packer. 1997. Habitat heterogeneity as a determinant of mammal species richness in high-energy areas. Nature 385: 252-254.
Lane, N. 2002. Oxygen; The Molecule That Made the World. Oxford University Press, Oxford.
Lovelock, J. E. 1988. The Ages of Gaia: A Biography of Our Living Earth. Norton, New York.
MacLeod, N. and G. Keller (eds.). 1996. Cretaceous-Tertiary Mass Extinctions: Biotic and Environmental Changes. W. W. Norton, New York.
Margulis, L. 1998. Symbiotic Planet: A New Look at Evolution. Basic Books, New York.
Maruyama, M. 1963. The second cybernetics: Deviation-amplifying mutual causal processes. American Scientist 51: 164-179.
McLean, D. M. 1978. A terminal Mesozoic "greenhouse": Lessons from the past. Science 201: 401-406.
Nicolis, G. and I. Prigogine. 1977. Self-organization in Nonequilibrium Systems. John Wiley, New York.
O'Brien, E. M. 1998. Water-energy dynamics, climate and prediction of woody plant species richness, an itnerim general model. Journal of Biogeography 25: 379-398.
Ostrom, J. 1969. Terrestrial vertebrates as indicators of Mesozoic climates. Proceedings of the North American Paleontological Convention: 347-376.
Paul, G. S. 2002. Dinosaurs of the Air: The Evolution and Loss of Flight in Dinosaurs and Birds. Johns Hopkins University Press, Baltimore.
Payton, A. and K. R. Arrigo. 2000. The sulfur-isotopic composition of Cenozoic seawater sulfate; implications for pyrite burial and atmospheric oxygen. International Geology Review 42: 491-498.
Powell, J. L. 1998. Night Comes to the Cretaceous: Dinosaur Extinction and the Transformation of Modern Geology. W. H. Freeman, New York.
Preston, F. W. 1948. The commonness and rarity of species. Ecology 29: 254-283.
Prigogine, I. 1961. Introduction to the Thermodynamics of Irreversible Processes. Interscience Publishers, New York.
Russell, D. A. 1982. The mass extinctions of the late Mesozoic. Scientific American 246: 48-55.
Schrodinger, E. 1945. What Is Life? Macmillan, New York.
Selas, V. 2006. UV-B induced plant stress as a possible cause of ten-year hare cycles. Population Ecology 48: 71-77.
Sinclair, A. R. E., D. Chitty, C. I. Stefan and C. J. Krebs. 2003. Mammal population cycles: evidence for intrinsic differences during snowshoe hare cycles. Canadian Journal of Zoology 81: 216-220.
Smith, C. H. 1983. Spatial trends in Canadian snowshoe hare, Lepus americanus, population cycles. Canadian Field-Naturalist 97: 151-160.
Smith, C. H. 1984. The Dynamics of Animal Distribution: An Evolutionary/Ecological Model. Ph.D. Dissertation, University of Illinois, Champaign-Urbana.
Smith, C. H. 1986. A contribution to the geographical interpretation of biological change. Acta Biotheoretica 35: 229-278.
Smith, C. H. 1989. Historical biogeography: geography as evolution, evolution as geography. New Zealand J. Zoology 16: 773-785.
Smith, C. H. 2004. Wallace's unfinished business. Complexity 10(2): 25-32.
Smith, C. H. 2005. Alfred Russel Wallace: past and future. J. Biogeography 32(9): 1509-1515.
Smith, C. H. and J. M. Davis. 1981. A spatial analysis of wildlife's ten-year cycle. J. Biogeography 8: 27-35.
Taylor, R. 1967. Causation. In Paul Edwards (ed.), The Encyclopedia of Philosophy. Macmillan, New York, Volume 2, pp. 56-66.
Vallyathan, V., X. Shi and V. Castranova. 2004. Oxygen/Nitrogen Radicals: Lung Injury and Disease. Marcel Dekker, New York.
Wiley, E. O., and D. R. Brooks. 1982. Victims of history--a nonequilibrium approach to evolution. Systematic Zoology 31: 1-24.
_________________________
Return
to Writings Menu
Return
to Home
Copyright 2006 by Charles H. Smith. All
rights reserved.
Materials from this site, whole or in part, may not be reposted or otherwise
reproduced for publication without the written consent of Charles H. Smith.
Feedback: charles.smith@wku.edu
http://people.wku.edu/charles.smith/once/finalc.htm