Historical
Biogeography:
Geography
as Evolution, Evolution as Geography
Charles H. Smith
[[Author's Note: An essay invited for publication
in the New Zealand Journal of Zoology (Volume 16: 773-785, 1989)
as part of a symposium issue, and reprinted here verbatim. Original pagination
indicated within double brackets.]]
[[p. 773]]
Abstract Despite a number of advances in method in recent
years, biogeography remains a field with a poorly developed philosophical
core. As a result, its historical and ecological sides remain as isolated
from one another as ever. In this essay I argue that a more unified approach
to biogeographic studies will become possible only when workers realise
that it is necessary to reject absolute space, "geography as handmaiden"
approaches to distribution problems in favour of structuralist models
compatible with both probabilistic spatial interaction and deterministic
phylogenetic kinds of thinking. Pros and cons of regionalist, vicariance,
and panbiogeographic approaches are weighed in this regard; it is shown
that the primary objections of the latter schools to the approach of the
former are vitiated when one dwells on second-order, rather than first-order,
interpretations of regional faunal structure. This approach makes it possible
to construct joint taxonomic/spatial models conducive to pattern analysis;
the latter permits the genesis of hypotheses that can be tested through
independently conceived theories of process (such as vicariance). An example
of the kind of pattern study envisioned, involving generalised track depiction,
is briefly described. A suggested cycle of research is thus laid out in
which systematic revision becomes a function of a joint "natural" spatial
and phylogenetic/historical approach to the subject.
Keywords panbiogeography; biogeography; vicariance; Leon Croizat; Alfred Russel Wallace
INTRODUCTION
Through this essay I should like to make a plea that one would not think necessary: to return the "geography" to (historical) biogeography. Between geographers and historical biogeographers there has been relatively little communication and, like Stoddart (1985), I consider this a problem. By far the majority of those publishing work with biogeographical themes are biologists. Most biology-trained biogeographers appear to have little or no familiarity with the theoretical, philosophical, and methodological literature of geography; this, at least, seems to be the only conclusion that can be drawn from the almost total absence of referral to such in their papers.
Geographers themselves are notorious "borrowers" of philosophy and theory from other fields, of course, but their applications and extensions of such are distinguished by the necessity of having to deal with the "how's"--not just "what's"--of very complex and explicitly spatial contexts. The overtly "open" nature of geographical systems has forced those who study related matters to specialise in the development and use of analytical techniques linking causal processes to spatial structure, and I feel it is time that biology-trained biogeographers became more conscious of these; and even more so, the philosophical rationale for their application.
Lest my arguments here be cast aside a priori as the irrelevant musings of a non-biologist, I should point out that there is some historical precedent for believing that biologists should give continuing attention to what might be called, for want of existing terminology, the "structural geography" of their subject. The two most important conceptual breakthroughs in the history of biogeography--those fashioned by Wallace and Croizat--consisted of new interpretations of the meaning of aggregate patterns of distribution, and this can hardly be viewed as other than a distinctly geographical matter.
Wallace is not generally recognised as a geographer (even, in fact, by geographers); elsewhere (Smith 1984a, unpubl. data), however, I have made what I feel is a strong argument that he was, in effect [[p. 774]] a geographer "with a consuming interest in the subject of evolution." As Fichman (1977) has shown, the faunal regions model he adopted strongly contributed to his overall evolutionary position: it provided a structure complementing his understanding of the relation of dispersal to biological change. Others (Brooks 1984; Browne 1983; McKinney 1972; Smith unpubl. data) have pointed out, moreover, that Wallace's views on species divergence and the theory of natural selection in general were closely tied to his interest in distribution-related matters.
[It perhaps should be emphasised at this point that Wallace did not fail "to recognise the fundamentals of biological classification"--as Nelson (1983, p. 489) has, by inference, accused. Wallace was no idle theoriser, being one of history's most productive field collectors, and the author of over 500 pages of systematic revisions (mostly of birds and insects). He wrote a fair amount on the theory of systematics as well, and a lot of what he had to say has a surprisingly modern feel (see, for example, Wallace 1856, 1863, 1864, 1874).]
Croizat, though primarily a biologist, also had a consuming interest in both distribution patterns per se and the geographical/ecological conditions contributing to these. Though he is sometimes viewed as an "anti-dispersalist," it is closer to the truth that his studies focused on exposing the limitations of dispersal as an explanatory vehicle. This paved the way, via Hennig's work (especially Hennig 1950, 1966), for the emergence of modern vicariance theory. Croizat's own synthesis, "panbiogeography", is notable for its rejection of a "standard-based" regional structuralism (i.e., the approach of Wallace and his followers) and promotion of a position emphasising taxon-specific trends of spatial affinity (i.e., the "generalised track" notion, which is usually logically extended to the study of inter-taxon trends as well).
In what follows I suggest that a "structural geography" approach--though not the one specifically adopted and promoted by Wallace--remains the most profitable starting point for historical biogeographic analysis. At the same time, I reject Wallace's dispersalism-dominated understanding of the evolution of present patterns of distribution, preferring Croizat's more adaptable "panbiogeography", which better lends its way to an integration of the role of vicariance. I hope to show, however, that the two approaches are not nearly so different as is now imagined, partly because Wallace's structural framework can be restated in more "natural" terms, and partly because many biologists have mistakenly (or tacitly) assumed that vicariant speciation events--quite possibly the primary immediate cause of distribution patterns--are synonymous with the process of evolution in general. As a result, I will also reject the attempts by panbiogeographers and vicariance biogeographers to make the "tree-thinking" of phylogeny-centred models the point of departure for spatial studies of evolution. Rather, I submit, it is more profitable to pursue taxon-level spatial analysis through: (1) an explicit consideration of the second-order characteristics of the overall distribution patterns of the group under study, followed by; (2) tests of theory suggested by such.
A GEOGRAPHER'S CRITICISM OF (HISTORICAL) BIOGEOGRAPHY AS A "BIOLOGICAL" STUDY
In a well-known paper, Gould & Lewontin (1979) produced a critique of the work of biologists who accept an "adaptations/adaptation" view of the nature of evolution by natural selection. I accept the majority of that discussion, which amounts to a plea that evolutionists not abandon structuralist interpretations of change linked to the "Bauplan" concept. Perhaps, however, the structural controls on evolution are to be more easily identified in facets of environmental organisation than in the individual lineages that trace their way through time as elements of the latter. This reasoning emerges from studies I have been carrying out in an attempt to realise a more internally consistent model of biological change than that offered by selectionist concepts.
Elsewhere (Smith 1984b, 1986c) I have worked out a theory of evolutionary change that focuses on the manner of turnover of resources vital to organismal existence; this, in contrast with models dwelling on organismal unit-level concepts such as "natural selection" and "competition" (nonetheless, characteristics of the latter processes are deducible from the process outlined in the model). To summarise a very long argument in a few words, I have suggested that range change can be viewed as a function of geographic variation in the characteristics of regularity in availability of, and optimality in level of, certain resources--especially water--vital to all life forms. The critical concept is that populations should probabilistically tend to extend toward areas where conditions are more optimum in the above sense, selecting into existence whatever new adaptive characteristics that "will [[p. 775]] work" as they do. Adaptation is thereby conceived as a correlative process of "keeping up with" the changes in spatial interaction implicit in organism-environment relations; i.e., an organism's adaptive suite may be considered a direct expression of all the interactions (and in turn morphological/physiological/behavioural adjustments) it must enter into to remain "at the right place at the right time", all the time, to collect/process all resources necessary to its existence.
From this starting point, a wide range of processes leading to observable characteristics of organismal distribution can be deduced; moreover, the perspective yields a fundamental criticism of "adaptation"-based thinking: the historical sequence of species divergence--and associated trends of adaptive change--define a conceptually different realisation of structure than does evolution in general. With respect to any given group (or all groups at once), the first process consists of an idiosyncratic response to environmental conditions yielding adaptive suites that mediate an ongoing ecological process of entropy maximisation. Adaptations are a resulting product of evolution but not, in the first instance, a cause of same. The character of a population's gene pool does, of course, constrain the range of possible further change in that lineage (in the sense of Wiley & Brooks 1982, 1986), but this restriction can be viewed as incidental to the evolutionary process as a whole, which more generally revolves around the creation of increasingly complex and efficient forms of interaction among organisms and their environment.
This construction completely resolves the "adaptation-process/adaptation-structure" dilemma stated by Lewontin (1984, pp. 237-238) as: "the process is adaptation and the end result is the state of being adapted . . . the problem is how species can be at all times both adapting and adapted." An organism can at one time be both "adapted" and "adapting" because both terms describe an ongoing process of change that can only be understood in terms of interactions between things. Strictly speaking, it is the conditions of interaction that "evolve", not the individual components that mediate this interaction. Thus, adaptations are epiphenomenal, reflecting (cryptic) standing relationships more than discrete structures. Viewed in this fashion, they constitute, in sum, mediators of an ongoing, probabilistically-realised resolution of environmental equilibrium.
This understanding leads to the conclusion that species populations should tend to expand or contract in common directions, though at rates peculiar to each (i.e., depending on the flexibility of the gene pool of each; for evidence, see Smith 1984b). This is significant for the possibilities it raises vis-a-vis normative forms of study: when the "individualistic hypothesis" is demonstrated in this fashion to be wholly fallacious (see Smith 1986c for discussion), distribution ranges can be treated as logically equivalent units of information, and their pattern of combination/correlation opened to probabilism-based interpretations.
The above concepts are related to the "Bauplan" matter as follows. As just suggested, what should most properly be considered as evolving on the surface of the earth is the surface of the earth--a complex pattern of spatial interaction among organisms and their environment (a concept not far removed from Croizat's definition of panbiogeography). This leads to the idea that none of the related concepts of divergence, adaptation, speciation, or specialisation represents a process that can, without introducing considerable illogic, be reasonably synonymised with the term "evolution". The four processes just listed are in effect evolutionarily neutral: any individual that comes into being eventually meets a finite end (whether through replacement, as in cells; death, as in individual organisms; extinction, as in populations, etc.). Existence in the form of populations, for example, is best interpreted as a response to ambient conditions that are prior; i.e., as a vehicle supporting a negative feedback loop that restores the equilibrium the overall system loses when species entities change range and must enter into new associations. As I interpret things, evolution per se is assured, as already suggested, by the tendency of populations to disperse toward less stressful environments (this (1) reduces the degree of specialisation required to exist, because the organism's energy budget can be apportioned to less stressful demands of "being in the right place. . ." type, and (2) eventually produces increasingly flexible and permanently appropriate populations whose interactions support a steady state resource turnover cycle--see discussion in Smith 1984b, 1986c). The point to be taken is that we would be better off looking for the causal basis of a "Bauplan" at the environmental--biogeographical--level than through examinations of secondary responses to the same (i.e., adaptive morphology and behaviour) that permit little in the way of generalisation (or even recognition, given the compounding difficulties of the idiosyncratic nature and usual obscurity of the effects themselves).
[[p. 776]] Organisms exist under probabilistic conditions of association with their environment (again, including other organisms). Although it is undoubtedly true that individual lineages represent stores of information (in the Wiley & Brooks sense, for example), such stores are little more than programmes for operation within a pre-existing setting. Though such programmes vary from taxon to taxon, their operation within their environment is maintained through probabilistic, not deterministic, interactions. Persistence as an element of an enveloping larger system is entirely a matter of information exchange. An organism is from top to bottom an open system; when it dies the locus of temporary storage it represented disappears, but is recapitulated both in its progeny and changes it effected in the composite nature of its environment (through its physical rearrangement of the "actors" contributing to the spatial interaction framework underlying the latter). It is in fact the patterns of interaction that maintain an "evolutionary progression": individual organisms (and populations thereof) may come and go, but such patterns must remain relatively constant or it will not be possible for any given individual that is born to fit into them. This brings us to a point of central importance to Bauplan-related investigations, the fact that the spatial interaction structure at the surface of the earth persists within a spatially-discrete domain, whereas the domain of adaptation is for all practical purposes unbounded.
I would argue that high on the list of priorities for biogeographical study should be a continuing search for normative studies-promoting frameworks. In this respect, an emphasis on spatial analytical concepts rather than character trait studies seems clearly preferable (speaking conceptually now, rather than methodologically, as will become clearer from later comment). The reason for this is fairly straightforward: spatial structure is more conducive to internally consistent forms of numerical measurement than are tree structures. I have already introduced the idea that it is possible to generalise on the meaning of pattern, on the basis of the understanding that organisms tend to change range in common directions. Beyond this, there is something even more fundamental: range change takes place within a domain that is absolutely delimited. That is, the logical complement to knowing where a population is found (in the sense of range) is knowing where it is not found, and vice versa. Presence/absence is a determinate and complementary condition spatially, and can be assigned relative probabilities on this basis. This is not true, of course, in the instance of character traits.
Now I grant that it is possible to understand characters as reflecting either an ongoing process of probabilistic interaction with environment or a causally determinate, historically-generated, sequence of relation to other groups of organisms (i.e., through patterns of common divergence). In the former instance, however, they are conceived as part of the environment; i.e., they function as loci through which information is exchanged, not stored. (Of course, storage does eventually take place via differential reproductive success/selection, etc., but this, as I am arguing, is a derivative, correlative process.) This primary function of adaptations is ignored when the populations they "reside within" are mapped onto a conceptually separate space and then treated as related to one another deterministically through some historical series of divergence. This is a classic example of application of an understanding that geographers have been fighting for many years: the "absolute space" notion of causality. Restricting "geography" and the "pattern of divergence" to conceptually different spaces leads us directly to the brink of environmental determinism; whenever we create artificially distinct spaces to sponsor single process understandings we are looking for trouble. Relaxing the constraints of reality in this fashion makes it all too easy to find what one wishes to find; although historical biology is not prone to the kinds of ethical lapses in application of theory the social sciences are in this regard, there is still the problem of thinking that the deterministic sequences of relation established, reflect an overall process of change reducible to such terms. Moreover, there is the difficulty that such formulations represent narratives often not amenable to test.
If, on the other hand, we allow our mappings of related populations to conceptually represent adaptive suites that mediate an environmental function, contexts for normative forms of study can be established. First, a single space is conceived, not distinct entities. Second, as spatial interaction can be interpreted: (1) as rates of change/process independent of the mediating structures, and (2) as a process that is fundamentally probabilistic in its enaction; it provides an internally consistent framework within which statistics-based methods can be used to isolate significant patterns of interaction. Further, the properties of interaction can be related to known sampling distributions, as the domain of interaction has explicit boundaries.
In another work (Smith 1984b) I have shown how deterministic pattern analysis methods can expose statistically significant trends of relation [[p. 777]] between range change and a posed environmental driving mechanism (note the deliberate use of the term "driving" rather than "determining"). Beyond such tests, there is also the possibility of simulation. On the assumption that range change events are probabilistically "driven" in preferred directions (as earlier described) toward more optimal environmental conditions, a diffusion process of range change (or pattern/rate of adaptive change) upon environment can be imagined. Thus, the study of spatial interaction can be reduced to purely spatial terms (involving prior assumptions/hypotheses about common directions--but differing rates--of diffusion among populations, the particular topological characteristics of the environmental "optimality surface" operating, etc.).
Although the above spatial conceptualisation of organic change does not directly characterise a "biogeography Bauplan", it suggests directions for further study. First, as a result of the bounded domain of the surface involved, the logical implication of operation of the mechanism is increasingly ordered interrelations, both among populations and between life and the inorganic sector of the environment. As I have discussed elsewhere (Smith 1984b), it may be possible to understand larger scale/longer term trends of biogeographic change on this basis.
Beyond the above, however, the fact that spatial interaction frameworks are measurable and referable to normative contexts can be exploited to provide methodological approaches that may be appropriate in a manner independent of scale of application. Later I will address this matter more directly; note, moreover, that it can be linked to arguments (which I will not expand on here: see Smith 1986a, b) that there actually may exist a controlling "biogeographical Bauplan" whose overt spatial expression is regional organisation.
With the preceding aside, let us briefly consider how the "speciation/organism" based schools of historical biogeography fall short of promoting spatially realistic biogeography interpretations. The oldest school, adhering to a regions-structured, dispersal-regulated interpretation, was popularised by Alfred Russel Wallace. Wallace considered the six-region classification system he adopted a "natural" one to the extent that it efficiently summarised what appeared to be a well-differentiated and general distributional response to underlying causal mechanisms. Nonetheless, in a number of respects it represents something less than the tag "natural" ought to reflect. To begin with, he never produced better than a correlative--history-based--understanding of these assumed mechanisms. Wallace adopted Sclater's (1858) original plan, which dealt with the distribution of birds. Finding it a satisfactory summary for the distribution patterns of mammals (which he considered the "model" group zoogeographically) he also concluded, as a package, that present distribution patterns were a function of dispersal and more or less permanently-located continental masses (Wallace 1877; Fichman 1977). But speciation has occurred at different rates in various times and places in various groups, and any attempt to create a general system based on any individual group is illogical, even if one accepts the "permanence" model (which itself, of course, turns out to be wrong). Moreover, the choice of six regions is very arbitrary; why not five, or seven, or whatever? Further, on what basis is each unit understood to be distinct? Unless it can be shown that all regions as established are of equal (structural) rank, of what use are they to comparative studies? From the point of view of spatial analysis, all these problems are highly relevant, because the use of probabilism-based methods assumes (as does, in effect, any kind of statistical analysis) the general equivalence and "naturalness" of the units under study.
Beyond the spatial systematics difficulties, moreover, there is the significant problem that Wallace's system is based on the concept that evolution proceeds principally through divergence following episodes of dispersal, a point of view that is now disputed. Further, world regional entities can only reasonably be recognised through the study of taxonomic units above the level of species populations. As should be apparent, I have no difficulty with the idea that evolution is primarily a function of dispersal; diversification per se, on the other hand, is quite another matter. Clearly, the ability of populations to expand range geographically is the fundamental spatial aspect of evolutionary change. Vicariance notwithstanding, we must initially get a population to a given location before any succeeding processes can operate. To ignore this idea is to ignore some rather fundamental things about the way a divergence-based process can operate in an actual spatial context. On the other hand, analysis of above-species-level patterns may obscure exactly what does happen once a population "gets there". Certainly, at the least the scale of inquiry is shifted from one in which immediate affinities (spatially, temporally, and phyletically, in a common origins sense) are emphasised to one in which more distant ones are.
[[p. 778]] There is finally the matter of the "natural" status of above-species level taxa and the implications of such for spatial classification. It seems to be just as difficult to argue that there is some set number of, for example, families, as it is to provide parallel rationale for a set number of regions. Implicit in this problem are the same kinds of effects on probabilism-based forms of analysis as those associated with "unnaturally-defined" regional units.
Through the work of Hennig (1950, 1966), Brundin (1966), Nelson (1969), and others, the structuralist biogeography of Wallace and followers was challenged and a new set of investigative methods introduced. The underlying concept, cladistics, provides a closely reasoned basis for the analytical retrieval of affinities (i.e., pattern of divergence) within a group of closely related populations.
The rigor of the concept, however, makes it difficult to apply to classification per se: any conditions of parallel evolution of traits subsequent to divergence are viewed as historically idiosyncratic, "unnatural", and no basis for grouping in a classificatory sense. This reasoning is in effect extended to the spatial arena; as a result, the biogeographical extrapolation of cladistics theory, vicariance analysis, consists, as mentioned earlier, of a literal projection of the history of the divergence patterns of the group under study onto a separately conceived, "geographic", space.
Despite the logical elegance of the cladistics concept and the apparent straightforward and parallel way it can be applied to historical studies, it represents a highly biased starting point for biogeographic investigations in general. Vicariance philosophy is utterly incapable of recognising the productive and functional role that spatial expansion processes have, including the formation of range conditions that make species divisions possible to begin with. Dispersal-based interactions are implicitly treated as instances in which "evolution has not taken place"; this, because it appears that many or most speciation episodes can be linked to intervening climatological and/or geological events which separate populations. But as I have suggested here and elsewhere (Smith 1984b, 1986c), evolution should not be considered a process synonymous with species divergence. Limited-ranging, highly specialised forms are (evolutionary) structure maintainers rather than structure innovators.
In another work (Smith unpubl. data) I have extended this argument to the point of showing how vicariant speciation can produce a return toward equilibrium when considered in spatial terms. In that study, which principally revolves around how to statistically expose generalised track relations, I show that a continuation from simple disjunct range patterns within a species to an outcome in which two new species are formed, results in a reduction in the overall statistical significance of the "track" connecting the locations involved. This finding is consistent with the earlier discussed idea that adaptive change per se is not evolutionary change; unless one argues that evolution can logically be interpreted as leading to generally lower levels of organisation (negentropy), it is difficult to view the spatial response indicated as falling within the class of deviation-amplifying processes, per Maruyama (1963). Maruyama (1963, p. 174) has also commented that when "the rules [of system evolution] are unknown, the amounts of information needed to discover the rules is much greater than the amount of information needed to describe the rules. This means that there is much more waste, in terms of the amount of information, in tracing the process backwards than in tracing it forward . . .", and in the philosophy posed by vicariance biogeographers one can recognise an instance in which this warning has gone unheeded.
Panbiogeography as a concept emerged earlier than did the vicariance biogeography school, originating with the studies of Leon Croizat. Although Croizat's work has been cited as instrumental in the development of the vicariance perspective (e.g., Nelson 1978), Croizat himself was careful to point out (for example, Croizat 1981) that the concepts were quite distinct. Panbiogeography has become popular as a method of study only in recent years, quite possibly as a function of an increasing recognition that he was quite right in this assessment. Conceptually, panbiogeography is flexible enough to allow that both dispersal and vicariance processes contribute to the production of distribution patterns; the main unifying element is the notion of the "generalised track", an observable trend of organismal affinities between geographically-separated locations which might be a function of the operation of either. With this core concept panbiogeography is distinctly more "geographic" in its approach than vicariance biogeography is: in theory, at least, it permits the adoption of relativistic analytical frameworks. Unfortunately, its practitioners also reject structuralist ("regionalist") interpretations of organismal geography, preferring the "absolute space" thinking of cladists as their base for unraveling spatial affinities (i.e., tracks). Craw (1988) has tried to associate geographers' "relative [[p. 779]] space" concept with the approach employed by cladists, but makes the fundamental error of failing to understand that the relative space concept is more properly defined in terms of real world interactions, not logical relations alone (as is the case in cladistics). This is a critical distinction; the geographer's primary chore, in fact, consists of assessing the degree to which the simplistic assumptions of closure implicit in aspatial logic fail to meet the reality of spatial organisation. (It should be no surprise that the fields of geography and psychology--which are, respectively, the most and least oriented toward study of "real world" spatial relationships--are afflicted, in turn, with the respective problems of having too little and too much independently standing theory. The constraints on existing as a three-dimensional reality are very strong). Cladistics (or panbiogeography, if it follows the lead of cladistics) cannot, even in principle, describe a geographically "relative space" until the objects of its analysis are re-defined into natural spatial terms; i.e., as a framework of spatially-induced probabilistic interactions.
Regardless of the above complaints, I believe that the three major competing schools of historical biogeography feature the following positive characteristics vital to the success of any synthesis of systematics based on compatible (natural) deterministic phylogenetic principles and (natural) probabilistic spatial interaction principles. Regionalisation approach/es (i.e., not necessarily Wallace's particular model) provide a basis for summarising in least biased fashion the overall process of evolution (as viewed biogeographically), and by virtue of their resting on interrelatable units are the best suited for the study of the spatial interaction process underlying the structure of these. The vicariance/cladistics-based approach rests on principles ensuring a sound basis for phylogenetic reconstruction and, therefore, classification (see later discussion); moreover, it provides one equally sound basis for testing/verifying hypotheses erected through the assessment of spatial interaction patterns (but not the reverse, as is now the way the matter is treated). Panbiogeography contributes a generalisable spatial focus for investigations of microprocess: situation-specific trends of spatial affinity within and/or across taxa, relatable to a variety of causal mechanisms.
Spatial-aspatial structuralism in biogeography
Though some might resist the notion a priori, I believe it is possible to harmonise the three main schools of historical biogeography in a fashion that respects the most desirable features of each of them individually. The key innovation involves the establishment of a second-order spatial analysis approach to distribution patterns that: (1) rests on structures specific to each monophyletic grouping of taxa, and (2) contributes to classification by producing feedback that introduces spatial criteria into the systematic revision process.
The easiest way to explain the approach I have in mind is to start with a description of the kind of interim results it produces. Let us first imagine we are handed a "natural" reconstruction of affinities within some monophyletic group. Our cladogram may actually be at any level of taxonomic organisation (see later discussion); for the present we will allow our base taxonomic unit to be the species. The geographic ranges of all species involved are mapped and the sum spatial domain of the group sampled; the "faunas" of each cell are then grouped into a spatial classification (preferably through the use of a non-hierarchical clustering algorithm, as these are better suited for this kind of partition problem). The result will be a first-order structure consisting of some limited number of spatial, "regional" units.
This descriptive device yields results that to one extent or another inherently reflect both the historical and ecological associations of the taxonomic units under study. Unless the underlying data are poor, moreover, the product represents a set of spatial units that are structurally very nearly (or absolutely) equivalent, rank-wise. It is little understood that the "non-equivalence" of regional units problem is, and has been all along, a pseudo-problem--a red herring. The criteria for rank assessment are quite different in spatial and aspatial contexts. Equivalence of rank in the spatial sense is a matter related to persisting balance of function, not equivalence of function. Persisting, "natural" structure may very well exist as a product of the sum of relations among non-equally unique subsystems; under such conditions the system is likely to be in a state of disequilibrium, but it seems pointless to argue that this means any given subsystem within it is more or less necessary to its overall coherence. First-order measures of relations between structural units tell us relatively little about the latter's "naturalness" as a group, because they are insensitive to the trade-off between structure and function. Before any progress can be made regarding the question of naturalness, the assumption of structural equality must be raised as a null hypothesis through some kind of entropy maximisation operation.
[[p. 780]] Wilson (1970) showed how it is possible to standardise the flows (specifically, commuter trips) among a set of source locations (in his case, a regional assemblage of towns and cities) in such a manner as to cancel out the effects of first-order system description induced by varying magnitude of flow (i.e., differing city populations). The operation involved, entropy maximisation, establishes a most-probable-state reformulation of within-system interaction. I have applied this approach to the study of biogeographical units (Smith 1983a, b), in the second work suggesting one way to test for functional rank equality (for another; see Smith 1986a, b; entropy maximisation procedures make possible the erection of the null hypothesis of structural/functional equality of units only--associated tests must rely on the introduction of additional assumptions).
Once the reliability of the spatial structure has been established (as discussed in Smith 1986a, unpubl. data), a second-order approach to regional faunal characterisation can be applied. Second-order statistics are probably best known for their contribution to nearest-neighbour analysis methods; my innovation has been to extend the concept in a fashion useful to the description of complex (overlapping) range patterns. These descriptive statistics can be used to very efficiently reconstruct the varying conditions of uniqueness exhibited by each fauna (Smith 1983b, c, unpubl. data). In a five-region model of world mammal differentiation patterns I have been working on, for example, the diversities of each regional fauna can be predicted via multiple regression from the first several vectors of second-order statistics (moments) describing each fauna to a level of efficiency reading r2 = .996. By contrast, my data for the Carnivora alone produced an analogous model yielding an r2 value of only .952. Carnivores, on the whole being adaptable generalists with large species ranges (Rapoport 1982), have not diversified in such a fashion as to produce well-defined regional units.
In the next section I will show how the above can be used as the basis for specific methodological approaches, but first it is necessary to sketch the overall framework into which such work can be placed.
Although I feel I have satisfactorily disposed of the "regional rank equivalence" problem, "organism-focused" biogeographers will still complain that a resolution to the "unnaturalness" of regional systemisation vis-a-vis organismal phylogeny and classification has not been suggested, and perhaps cannot be. I believe, however, that it can.
Regardless of my adherence to a structuralist regional approach, I fully support cladistics-based efforts at unraveling phylogeny. For any effort at spatial classification to succeed, we must know that the group is monophyletic. This makes it easier to argue that there are logical limits to the group's spatial domain. The group need not be restricted to species-level (taxonomic and spatial) units, however. Recalling comments made earlier, I suggest that classification should devolve as a function of a combined understanding of spatial patterns and the aspatial reconstruction of phylogeny. As in the case of regional rank assessments, taxonomic rank equivalence at the above-species level must be viewed as a matter of function as well as affinities. There is no compelling reason why family units, for example, need be "equivalent" in terms of their first-order characteristics of uniqueness (including their varying numbers of species). Further, it does not conflict with the idea of natural classification to accept the existence of paraphyletic family units. As I suggested in Smith (1986b), "naturalness" of relation is a combined function of historical derivation and environmental function. Organisms are, in the first instance, environmental units, not historical ones. We are yet a long way from understanding to what degree genetic differences are expressed as operationally different adaptive strategies at the ecological level, and to rely on "degree of structural difference" arguments as the basis for understanding the "natural" degree of distinctness among populations is to subscribe to a very tenuous definition of "natural". What are, relatively speaking, only minor genetic differences can have profound ramifications in terms of function (consider the case of Pan and Homo, for example). Not surprisingly, biochemistry- and morphology-based analyses of "natural relations" often produce conflicting results: spatial form can never be interpreted in wholly consistent fashion in terms of selected one-to-one mappings of divergence on a prior space. Taxonomic classification per se, I submit, should represent as rational and accurate an accounting as possible of this trade-off between historical inertia and current environmental function; as such, it should take into account all kinds of information relevant to establishing systematic position.
Biogeographic studies can play a major role in such efforts. If one assumes a probabilistic model of the resolution of genetic potential and environmental constraint/opportunity (as sketched earlier, for example), the overall spatial pattern of differentiation of a given group can be used as a working hypothesis
[[p. 781]]
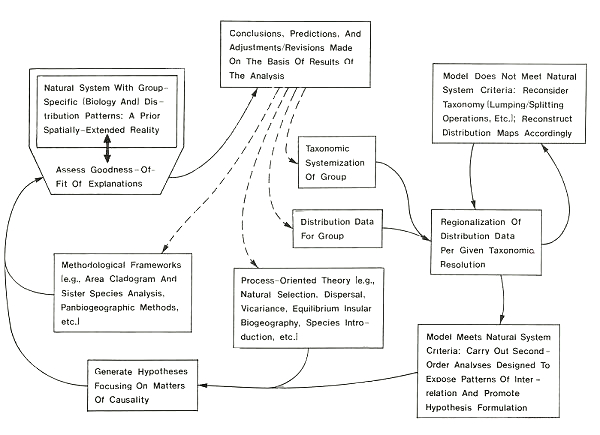
Fig. 1. A cycle of biogeographic research suggested by the "natural system" concepts introduced in Smith (1986a, b). The purpose of this cycle is to establish a unified kind of approach to the study of the spatial and aspatial systematics of any taxon under study. See text for discussion.
regarding the classification (i.e., taxonomic) status of each unit initially assumed for purposes of organising range data. In Smith (1986b, unpubl. data) I describe methods whereby a regionalisation of initial distribution data that cannot be validated (i.e., in which the regional units determined fail to meet the rank equivalence criteria making such decisions possible) can be revised in such a manner as to provide feedback for taxonomic revision. Lumping and splitting operations are initiated on those taxa whose status is most debated until a spatial resolution of the resulting set of range data is obtained that can be validated. The result is a hypothesis of structure that can then be described via pattern analysis and subjected to test in relation to process models. In Fig. 1, the essence of this procedural cycle is represented.
The cycle of analytical input described through Fig. 1 has no specific beginning, but is continually self-refining. One of the important implications of the philosophical framework underlying it (as discussed in Smith, 1986a, b) is the concept that a "natural" taxonomic resolution must have a parallel "natural" spatial resolution; i.e., the former can always be linked to a "most-probable-state"-based interpretation of actual spatially-extending structure. When through an analysis of appropriate data this cannot be verified, both aspects of the systematics model must be re-assessed and adjusted until reciprocally valid. Tests of the resulting hypothesis are then constructed through pattern analysis and relation to independently existing process models. When the model as tested does not hold up, the implications are reconsideration of its fundamental elements--especially the underlying taxonomic resolution (e.g., is the group actually monophyletic?) and the adequacy (generality) of the causal models that have been brought to bear in its interpretation.
Note that the entire cycle described above can be implemented strictly on the basis of null hypotheses revolving around most-probable-state concepts. In the spatial sense, hypothesis tests are grounded in [[p. 782]] the idea that once the most-probable-state of the internal structure of the system has been identified, measured deviations from this (its second-order structure) can be interpreted as indicative of the presence of external (environmental/historical) causal mechanisms. Otherwise put, once we have satisfactorily isolated the structural components underlying first-order patterns of faunal differentiation, tests may be applied on this basis to determine the extent to which the second-order relations exposed fail to fall within statistically probable assumptions (null hypotheses) about random patterns of interaction. Two examples of such kinds of study can now be briefly described.
The methodological application of second-order concepts
I have described a method for the study of generalised tracks in some detail in another work (Smith unpubl. data). In past years the main criticism of the "generalised track" concept has been its apparent resistance to statistical studies (McDowall 1978; Simberloff et al. 1981). The essence of the criticism is that the non-random trends within a uniquely-defined system are difficult to distinguish from conditions of randomly defined association. Attempts have been made to apply randomisation methods (Simberloff 1978; Simberloff & Connor 1979; Connor & Simberloff 1983) and graph theory (Craw & Page 1988) to related ecological and historical aspects, respectively, of the problem.
The ecological approaches are beyond the scope of present discussion; the graph theory approach proposed by Page (1987) and Craw & Page (1988) seems to require a priori assumptions about the relation of absolute distance to biological affinities that it would be better to eliminate. It is more efficient to incorporate ideas concerning these into investigative frameworks as hypotheses than as assumptions; i.e., there are few such links that are of such generality as to constitute laws of relation. There are, moreover, the problems of how to accommodate large data sets through this approach and produce results convenient to consensus studies.
My own treatment of the matter is based on the earlier discussed strategy of determining the first-order structure (i.e., regional units) of the group pattern of distribution of the taxon under study, and using this as the basis for a second-order analysis through which improbable conditions of spatial affinity between the faunas of particular locations are identified. The method is complicated enough that it cannot be described in detail here; the interested reader is referred to the other work (Smith unpubl. data). The general approach involved can, however, be sketched. (Note that what follows is associated largely with the "model meets natural system criteria" box in Fig. 1).
Once a regionalisation model consistent with the taxonomic data has been established, "first-order" (presence-absence) similarity scores are calculated for the faunal relations between each pairing of cells in the sampling grid (see Smith 1983b, unpubl. data, for discussions of appropriate derivations). A pair of parallel sets of second-order calculations are then performed. These scores are formulated in such a manner as to represent entropy maximisations of the first-order scores; i.e., a standardisation procedure is applied to the first-order relations which explicitly takes into account the varying cosmopolitanism of each taxonomic unit. For each cell's relations with all other cells, regression models are then set up: the test hypothesis is that residuals from a regression of the first-order scores on the second-order scores should not be correlated with the dependent variable (i.e., if there is no non-random component to the second-order influence on first-order structure). Otherwise put, an attempt is made to determine whether we can reject the assumption that the overall pattern of overlap of ranges can be interpreted as deviating from a simple most-probable-state kind of spatial organisation.
Each cell is treated in this manner, and is thus assigned; (1) an r value for each original regression, and (2) a pair of r values for the correlations of the residuals produced back on the first-order scores. Two sets of r values are obtained in #2 above because two different sets of second-order scores are calculated for each cell's affinities: one focusing on the pattern of "presences" of all taxa, and an entirely complementary one focusing on "absences" of same. (Recall that this is logically appropriate for binary characteristics existing as part of an actual spatial domain, in contrast with the situation in character analysis.) In effect, two "one-tailed" tests are thus obtained: one focusing on improbable faunal relations in terms of widespread forms, and one on same for forms whose ranges are more restricted. The greater the violation of the assumption of independence of cases, the more substantial the "preferred orientation" of other cells with respect to the base cell. The complementary models may be interpreted, as a first approximation, in vicariance and dispersal terms, respectively. It is often the case, as might be expected, that only one of the two models exhibits significantly non-random patterns of affinity. It is quite possible, [[p. 783]] however, for both (or neither, or course) to do so. It should be emphasised that the r scores associated with each cell's models are in every sense logically and numerically compatible as measurements; this is ensured by the second-order approach employed (see Smith 1983c, unpubl. data).
Each set of residuals can then be assessed as desired for its elements' properties of spatial autocorrelation. Regardless of the overall degree of "preferred orientation", high residual values will be indicative of improbable degrees of association (which may be causally incidental, of course--further investigation is then necessary to ensure a correct interpretation). At this point, as can be seen from Fig. 1, causal hypotheses for the non-random patterns identified can be entertained.
Through this approach, the characteristics of spatial affinity between any pair of locations bearing on any given set of data can be statistically investigated. Moreover, as suggested in Smith (unpubl. data), new kinds of questions regarding biogeographic and evolutionary dynamics can be posed and studied through it. For example, the extent to which vicariance represents a general process of return toward system equilibrium (i.e., the "devolution" concept commented on earlier) rather than a deviation-amplifying process might be explored. (Again, it turns out that vicariant events can actually reduce degree of violation of the assumption of unconditional independence of cases). Another useful feature is that trends involving very large groups are especially amenable to this method; unlike phylogeny-based studies (note Simberloff 1987), in fact, the more taxa, the better.
Nevertheless, I see no reason why the same approach cannot be applied to small groups, or even a single population, if the focus is the geographic distribution of character traits rather than population distribution per se. The concept behind the method is scale-independent; there must, however, be sufficient spatial interaction implied among the initial biological (i.e., taxonomic or character) units represented to permit the establishment of a meaningful most-probable-state standard. It should be possible to establish gene flow/variation studies within a given population in directly analogous fashion to supra-species level biogeographic investigations. Subspecies should partition out in a manner paralleling the isolation of regional structures. "Track" investigations would focus on possible causal mechanisms, whether these be climatic, biological (e.g., a function of parallel evolution, commensalism/parasitism, competition, etc.), environmental (protective coloration, climate, etc.), or whatever. It even seems that geographic variation studies on, for example, a single wide-ranging member of a large group might be used to confirm parallel studies on the overall group--especially as distribution patterns within the group, can be linked to varying ambient environmental influence (e.g., clines).
CONCLUSION
The preceding ideas might initially strike one as deviating considerably from the subject of the symposium dealt with in this issue, but the deviation may be more apparent than real. Panbiogeography, as realised by Croizat, at heart, it seems to me, represented an effort to draw attention to: (1) possible non-dispersal-based interpretations of the evolutionary process, and (2) the importance of giving greater attention to unique patterns defined in terms of organisms, not "prior" (and untestable) structuralist concepts. Current practitioners appear to have adopted an entirely organismal approach, not realising that a general spatial framework for evolutionary distribution studies based directly on an application of cladistic principles is, even in theory, not possible. The "geography as handmaiden" approach to biogeography is a contradiction in terms: the distribution of organisms is not assignable to a (prior) geographical space, it is geographical space.
I am therefore confident that biogeography--whether treated in historical or ecological terms--is to be more efficiently dealt with through probabilism-based modelling approaches. I am also confident that it will come to be recognised that dispersal, rather than vicariance, represents the dominating process of "inventive" evolution (i.e., that which leads as a progression to organisms and environmental associations of higher structural complexity). The confusion on this issue arises from the unfortunate oversimplication that adaptation per se, and thus divergence, is synonymous with evolution. I submit instead that in the most part divergence represents a return toward, rather than movement away from, equilibrium. The main feedback produced by range change--and entrance into new conditions of spatial interaction--is adaptation; the significance of divergence is the way adaptive suites can be refocused to react to local conditions of environmental constraint on efficiency of resource turnover. The more efficient the turnover process, the less constraining the environment becomes; thus is prepared an ever more stable base and the possibility of very highly "improbable" organic structure. We do not [[p. 784]] need teleology to support a "Gaic hypothesis" interpretation of world ecosystem change: probabilistic interaction (i.e., spatial interaction) within a limited spatial domain naturally leads to both increasing order and increasing diversity.
The comments offered here may be viewed as criticisms of present strategies of biogeographic investigation, but this is not the whole story. The gap between the panbiogeography school and concepts I have discussed here is actually rather small. I envision a regional structural approach defined by the interaction patterns of logically-constituted organismal taxonomic units, and peculiar to each such group. I cannot accept the naive understanding that a historically-defined phylogeny--a set of logical relations that in of itself carries no capacity for external validation--is in some sense more "natural" than ecologically organised reality. This does not mean, however, that the two understandings cannot be integrated within a position recognising the contributions of both to making the reality comprehensible. As discussed earlier and in Smith (1986a, 1986b), it is quite possible to recognise a normative structural basis for the interpretation of unique systems of organisation; Croizat's objections to the structural interpretation of the regionalists are thereby rendered irrelevant.
But this is no more than we should expect. Croizat's attempt to bury Darwin and Wallace was, as is often the case when contradictory discoveries are first made, an overreaction. The leading result of his work, the generalised track, is itself a structuralist concept. Spatial organisation is all structure. History is nothing beyond the selective interpretation of changing structure. (Regional geography and history have been closely associated as studies since ancient times, and this can hardly be viewed as surprising in this light.) The idea that a "complete" understanding of the nature of (any kind of) spatial organisation can be obtained by partitioning the observable elements of the latter into independently-defined entities is a conceptual dinosaur.
Panbiogeography cannot live up to the "pan" part of its name until its practitioners return the "geography" element of their charge to an appropriate role.
REFERENCES
Brooks, J. L., 1984: Just before the origin: Alfred Russel Wallace's theory of evolution. New York, Columbia University Press.
Browne, J., 1983: The secular ark; studies in the history of biogeography. New Haven & London, Yale Univ. Press.
Brundin, L., 1966: Transantarctic relationships and their significance, as evidenced by chironomid midges. Kungliga Svenska Vetenskapsakademiens Handlingar 2 (4th ser.) (1).
Connor, E. F.; Simberloff, D., 1983: Interspecific competition and species co-occurrence patterns on islands: null models and the evaluation of evidence. Oikos 41: 455-465.
Craw, R. C., 1988: Panbiogeography: method and synthesis in biogeography. In: Myers, A. A.; Giller, P. S., ed., Analytical biogeography. London & New York, Chapman & Hall. pp. 405-435.
Craw, R. C.; Page, R., 1988: Panbiogeography: method and metaphor in the new biogeography. In: Ho, M. -W.; Fox, S., ed., Evolutionary processes and metaphors. New York, etc., John Wiley & Sons. pp. 163-189.
Croizat, L., 1981: Biogeography: past, present, and future. In: Nelson, G.; Rosen, D. E., ed., Vicariance biogeography: a critique. New York, Columbia Univ. Press. pp. 501-523.
Fichman, M., 1977: Wallace: zoogeography and the problem of land bridges. Journal of the history of biology 10: 45-63.
Gould, S. J.; Lewontin, R. C., 1979: The spandrels of San Marco and the Panglossian paradigm: a critique of the adaptationist programme. Proceedings of the Royal Society, London 205B: 581-598.
Hennig, W., 1950: Grundzuge einer Theorie der phylogenetischen
Systematik. Berlin, Deutscher Zentralverlag.
_____1966: Phylogenetic systematics. Urbana, University
of Illinois Press.
Lewontin, R. C., 1984: Adaptation. In: Sober, E., ed., Conceptual issues in evolutionary biology. Cambridge, MIT Press. pp. 235-251.
McDowall, R. M., 1978: Generalised tracks and dispersal in biogeography. Systematic zoology 27: 88-104.
McKinney, H. L., 1972: Wallace and natural selection. New Haven, Yale University Press.
Maruyama, M., 1963: The second cybernetics: deviation-amplifying mutual causal processes. American scientist 51: 164-179.
Nelson, G., 1969: The problem of historical biogeography.
Systematic zoology 18: 243-246.
_____1978: From Candolle to Croizat: comments on
the history of biogeography. Journal of the history of biology 11:
269-305.
_____1983: Vicariance and cladistics: historical
perspectives with implications for the future. In: Sims, R. W.;
Price, J. H.; Whalley, P. E. S. ed., Evolution, time and space:
the emergence of the biosphere. London, etc., Academic Press. pp. 469-492.
[[p. 785]] Page, R. D. M., 1987: Graphs and generalised tracks: quantifying Croizat's panbiogeography. Systematic zoology 36: 1-17.
Rapoport, E. H., 1982: Areography; geographical strategies of species. Oxford, Pergamon Press.
Sclater, P. L., 1858: On the general geographical distribution of the members of the class Aves. Proceedings of the Linnean Society, London 2: 130-145.
Simberloff, D., 1978: Using island biogeographic distributions
to determine if colonisation is stochastic. American naturalist 112:
713-726.
_____1987: Calculating probabilities that cladograms
match: a method of biogeographical inference. Systematic zoology 36:
175-195.
Simberloff, D.; Connor, E. F., 1979: Q-mode and R-mode analyses of biogeographic distributions: null hypotheses based on random colonisation. In: Patil, G. P.; Rosenzweig, M. L., ed., Contemporary quantitative ecology and related econometrics. Fairland, Maryland, International Co-operative Publishing House. pp. 123-138.
Simberloff, D.; Heck, K. L.; McCoy, E. D.; Connor, E. F., 1981: There have been no statistical tests of cladistic biogeographical hypotheses. In: Nelson, G.; Rosen, D. E., ed., Vicariance biogeography: a critique. New York, Columbia University Press. pp. 40-72.
Smith, C. H., 1983a: A system of world mammal faunal
regions. I. Logical and statistical derivation of the regions. Journal
of biogeography 10: 455-466.
_____1983b: A system of world mammal faunal regions.
II. The distance decay effect upon interregional affinities. Journal
of biogeography 10: 467-482.
_____1983c: Areographic representation of faunal
characteristics through a 'second order' relational approach. Evolutionary
theory 6: 225-232.
_____1984a: Alfred Russel Wallace. In:
Freeman, T. W., ed., Geographers: biobibliographical studies,
London, Mansell, vol. 8. pp. 125-133.
_____1984b: The dynamics of animal distribution:
an evolutionary/ecological model. Unpublished PhD. Dissertation, Univ.
of Illinois, U.S.A.
_____1986a: A general approach to the study of
spatial systems. I. The relational representation of measurable attributes.
International journal of general systems 12: 359-384.
_____1986b: A general approach to the study of
spatial systems. II. Two examples of application. International journal
of general systems 12: 385-400.
_____1986c: A contribution to the geographical
interpretation of biological change. Acta biotheoretica (Leiden) 35:
229-278.
Stoddart, D. R., 1985: On geography. London, Basil Blackwell.
Wallace, A. R., 1856: Attempts at a natural arrangement
of birds. Annals and magazine of natural history 18 (2nd s.):
193-216.
_____1863: Who are the humming bird's relations?
Zoologist 21: 8486-8491.
_____1864: Remarks on the value of osteological
characters in the classification of birds. Ibis 6: 36-41.
_____1874: On the arrangement of the families constituting
the order Passeres. Ibis 4 (3rd s.): 406-416.
_____1877: The comparative antiquity of continents,
as indicated by the distribution of living and extinct animals. Proceedings
of the Royal Geographical Society 21: 505-534.
Wiley, E. O.; Brooks, D. R., 1982: Victims of history:
a nonequilibrium approach to evolution. Systematic zoology 31:
1-24.
_____1986: Evolution as entropy. Chicago &
London, University of Chicago Press.
Wilson, A. G., 1970: Entropy in urban and regional planning. London, Pion Press.
*
*
*
*
*
Copyright 1989, 2002 by Charles H. Smith. All rights reserved.